Hebbisk teori
On december 14, 2021 by admin Bedömning | Biopsykologi | Jämförande | Kognitiv | Utveckling | Språk | Individuella skillnader | Personlighet | Filosofi | Social |
Metoder | Statistik | Klinisk | Pedagogisk | Industriell | Professionella artiklar |Världspsykologi |
Kognitiv psykologi:Uppmärksamhet – Beslutsfattande – Inlärning – Bedömning – Minne – Motivation – Perception – Resonemang – Tänkande – Kognitiva processerKognition -OutlineIndex
Hebbianska teorin beskriver en grundläggande mekanism för synaptisk plasticitet där en ökning av den synaptiska effektiviteten uppstår genom den presynaptiska cellens upprepade och ihållande stimulering av den postsynaptiska cellen. Den introducerades av Donald Hebb 1949 och kallas också Hebbs regel, Hebbs postulat och cellsamlingsteori, och lyder:
Låt oss anta att persistensen eller upprepningen av en reverbererande aktivitet (eller ”spår”) tenderar att framkalla varaktiga cellförändringar som bidrar till dess stabilitet…. När en axon från cell A är tillräckligt nära för att excitera en cell B och upprepade gånger eller ihållande deltar i avfyrningen av den, sker någon tillväxtprocess eller metabolisk förändring i en eller båda cellerna så att A:s effektivitet, som en av de celler som avfyrar B, ökar.
Teorin sammanfattas ofta som ”celler som eldar tillsammans, kopplar ihop”, även om detta är en överdriven förenkling av nervsystemet som inte ska tas bokstavligt, samt att det inte korrekt återger Hebbs ursprungliga uttalande om förändringar i styrka av cellkopplingar. Teorin används ofta för att förklara vissa typer av associativ inlärning där samtidig aktivering av celler leder till en kraftig ökning av den synaptiska styrkan. Sådan inlärning kallas hebbisk inlärning.
Hebbiska engram och cellsamlingsteori
Hebbs teori handlar om hur neuroner kan koppla samman sig själva för att bli engram. Hebbs teorier om cellföreningars form och funktion kan förstås av följande:
”Den allmänna idén är gammal, att två celler eller system av celler som upprepade gånger är aktiva samtidigt tenderar att bli ’associerade’, så att aktivitet i den ena underlättar aktivitet i den andra”. (Hebb 1949, s. 70) ”När en cell upprepade gånger hjälper till med att tända en annan, utvecklar den första cellens axon synaptiska knoppar (eller förstorar dem om de redan finns) i kontakt med den andra cellens soma”. (Hebb 1949, s. 63)
Gordon Allport lägger fram ytterligare idéer om teorin om cellförsamling och dess roll i bildandet av engram, i linje med begreppet autoassociation, som beskrivs på följande sätt:
”Om ingångarna till ett system gör att samma aktivitetsmönster uppträder upprepade gånger, kommer uppsättningen av aktiva element som utgör det mönstret att bli alltmer starkt interassocierade. Det vill säga, varje element kommer att ha en tendens att aktivera varje annat element och (med negativa vikter) att stänga av de element som inte ingår i mönstret. För att uttrycka det på ett annat sätt kommer mönstret som helhet att bli ”autoassocierat”. Vi kan kalla ett inlärt (autoassocierat) mönster för ett engram.” (Hebb 1949, s. 44)
Hebbianska teorin har varit den främsta grunden för den konventionella uppfattningen att engram, när de analyseras från en holistisk nivå, är neuronala nät eller neurala nätverk.
Arbete i Eric Kandels laboratorium har gett belägg för att hebbiska inlärningsmekanismer är inblandade i synapser hos havssnäckan Aplysia californica.
Experiment om Hebbiska synapsmodifieringsmekanismer vid synapserna i det centrala nervsystemet hos ryggradsdjur är mycket svårare att kontrollera än experiment med de relativt enkla synapserna i det perifera nervsystemet som studeras hos marina ryggradslösa djur. En stor del av arbetet med långvariga synaptiska förändringar mellan neuroner hos ryggradsdjur (t.ex. långtidspotentiering) innebär att man använder sig av icke-fysiologisk experimentell stimulering av hjärnceller. Några av de fysiologiskt relevanta synapsförändringsmekanismer som har studerats i ryggradsdjurshjärnor verkar dock vara exempel på hebbiska processer. En sådan studie går igenom resultat från experiment som visar att långvariga förändringar i synaptiska styrkor kan induceras av fysiologiskt relevant synaptisk aktivitet som fungerar genom både hebbiska och icke-hebbiska mekanismer
Principer
Från synvinkeln av artificiella neuroner och artificiella neurala nätverk kan Hebb-principen beskrivas som en metod för att bestämma hur man ändrar vikterna mellan modellneuroner. Vikten mellan två neuroner ökar om de två neuronerna aktiveras samtidigt – och minskar om de aktiveras separat. Noder som tenderar att vara antingen båda positiva eller båda negativa samtidigt har starka positiva vikter, medan de som tenderar att vara motsatta har starka negativa vikter.
Denna ursprungliga princip är kanske den enklaste formen av viktval. Även om detta innebär att den relativt enkelt kan kodas in i ett datorprogram och användas för att uppdatera vikterna för ett nätverk, förbjuder det också antalet tillämpningar av hebbisk inlärning. I dag hänvisar termen hebbisk inlärning i allmänhet till någon form av matematisk abstraktion av den ursprungliga princip som Hebb föreslog. I denna mening innebär hebbisk inlärning att vikterna mellan inlärningsnoderna justeras så att varje vikt bättre representerar förhållandet mellan noderna. Många inlärningsmetoder kan därför anses vara något Hebbiska till sin natur.
Nedan följer en formelmässig beskrivning av Hebbisk inlärning: (Observera att många andra beskrivningar är möjliga)
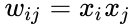
där 
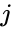
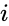
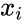
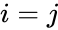
En annan formelbeskrivning är:
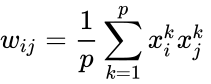
där 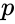

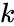
En variant av hebbisk inlärning som tar hänsyn till fenomen som blockering och många andra neurala inlärningsfenomen är Harry Klopfs matematiska modell. Klopfs modell reproducerar ett stort antal biologiska fenomen och är dessutom enkel att implementera.
Generalisering och stabilitet
Hebbs regel generaliseras ofta som
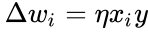
eller förändringen i den 
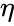
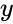
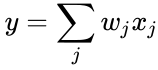
och i föregående avsnitts förenkling antas både inlärningshastigheten och ingångsvikterna vara 1. Denna version av regeln är helt klart instabil, eftersom i alla nätverk med en dominerande signal kommer de synaptiska vikterna att öka eller minska exponentiellt. Det kan dock visas att Hebbs regel är instabil för vilken neuronmodell som helst. Därför används i nätverksmodeller av neuroner vanligen andra inlärningsteorier som BCM-teorin, Ojas regel eller den generaliserade hebbiska algoritmen.
Se även
- Anti-Hebbs inlärning
- BCM-teori
- Koincidensdetektion i neurobiologi
- Dales princip
- Generaliserad Hebbisk Algoritm
- Leabra
- Long-term potentiation
- Minne
- Metaplasticitet
- Neurala nätverk
- Oja inlärningsregel
- Tetanisk stimulering
- Spike-timing-beroende plasticitet
- Synaptotropisk hypotes
- Shouval, Harel Hjärnans fysik. Den synaptiska grunden för inlärning och minne: Ett teoretiskt tillvägagångssätt. University of Texas Health Science Center i Houston. URL tillgänglig 2007-11-14.
Vidare läsning
- Hebb, D.O. (1949), The organization of behavior, New York: Wiley
- Hebb, D.O. (1961). ”Distinctive features of learning in the higher animal” J. F. Delafresnaye (Ed.) Brain Mechanisms and Learning, London: Oxford University Press.
- Hebb, D.O., and Penfield, W. (1940). Mänskligt beteende efter omfattande bilateralt avlägsnande från frontalloberna. Archives of Neurology and Psychiatry 44: 421-436.
- Allport, D.A. (1985). ”Distributed memory, modular systems and dysphasia” Newman, S.K. and Epstein, R. (Eds.) Current Perspectives in Dysphasia, Edinburgh: Churchill Livingstone.
- Bishop, C.M. (1995). Neural Networks for Pattern Recognition, Oxford: Oxford University Press.
- Paulsen, O., Sejnowski, T. J. (2000). Naturliga aktivitetsmönster och långsiktig synaptisk plasticitet. Current opinion in neurobiology 10 (2): 172-179.
- Översikt
- Hebbian Learning tutorial (Del 1: Novelty Filtering, Del 2: PCA)
|
v-d-e
Neuroetologi |
||
|---|---|---|
| Begrepp inom neuroetologi |
Feedforward – Coincidence detector – Umwelt – Instinkt – Funktiondetektor – Central mönstergenerator (CPG) -NMDA-receptor – Lateral hämning – Fast handlingsmönster – Kroghs princip-Hebbianska teorin – Ljudlokalisering |
200px
|
| Neoetologins historia |
Theodore Holmes Bullock – Walter Heiligenberg -Niko Tinbergen- Konrad Lorenz- Eric Knudsen-Donald Griffin – Donald Kennedy – Karl von Frisch – Erich von Holst – Jörg-Peter Ewert |
|
| Metoder inom neuroetologin |
Helcells-patchklämma |
|
| Modellsystem inom neuroetologin |
Djurens ekolokalisering – vickning Dance- Electric Fish – Vision in toads – Neuroethology of Frog Audition – Infrared sensing in snakes |
|
Denna sida använder sig av Creative Commons licensierat innehåll från Wikipedia (visa författare).
Lämna ett svar