Frontiers in Psychology
On september 28, 2021 by adminCorpus callosum (CC) är den primära kommissuren som förbinder de två hjärnhalvorna i hjärnan. Även om termen commissurotomi inte är felaktig för att beskriva den så kallade ”delade hjärnan”, och är strikt korrekt för de ursprungliga kirurgiska ingreppen på människor där alla commissurer skars av (Gazzaniga et al, 1967), så skar senare operationer bara av CC, vilket ledde till att termen ”callosotomi” började användas, som förblir den mer lämpliga termen i dessa fall (t.ex. Franz et al., 1996, 2000).
Från ett evolutionärt perspektiv är CC en ny utveckling som endast observerats hos däggdjur med placenta (Katz et al., 1983; Mihrshahi, 2006). Den funktionalitet som tillhandahålls av CC tros ha föregåtts, åtminstone delvis, av den främre kommissuren (AC), som observerats hos icke-placentala däggdjur, t.ex. pungdjur (Heath och Jones, 1971; Ashwell et al., 1996). Även om vissa återstående vägar från AC fortfarande finns kvar i den mänskliga hjärnan, är dess konnektivitet och struktur fortfarande understuderad i den aktuella litteraturen och de få resultaten är ganska inkonsekventa (Lasco et al., 2002; Patel et al., 2010; Choi et al., 2011). Till exempel är resultaten baserade på magnetisk resonanstomografi (MRI) blandade när det gäller huruvida AC är lika stor hos kvinnor och män (Lasco et al., 2002; Patel et al., 2010). I en relativt ny studie där man använde sig av in-vivo-tekniken diffusionstensorimaging (DTI), som utnyttjar spridningen av vattenmolekyler i axoner, studerade Patel et al. (2010) åtta försökspersoner och lyckades isolera fibrer från AC hos fem försökspersoner. Bland fynden fanns hos en försöksperson en stor andel fibrer i AC:s bakre extremitet som rörde sig bilateralt in i parietala regioner där de flesta av dem slutade, med ett mindre knippe som slutade i gyrus precentral och ytterligare ett knippe som gick in i temporalloben. Sådana individuella skillnader hittades hos alla deltagare men ändå visade majoriteten av försökspersonerna bilaterala projektioner till de orbitofrontala, parietala och temporala kortiklarna, av vilka CC inte tydligt projicerar till. Tillsammans innebär resultaten att AC har både primära och rudimentära projektioner till neokortikala områden där CC inte projicerar, vilket andra också har föreslagit (Di Virgilio et al., 1999; Patel et al., 2010).
Enligt vår åsikt förlitar sig AAA i hög grad på neokortikala nätverk och konnektivitet med subkortikala strukturer som basala ganglier och thalamus (bland annat andra som inte diskuteras specifikt i vårt förslag) (Franz, 2012). Alltså kan en eventuell evolutionär utveckling som involverar AC också ha en viss roll i AAA-modellen. Vi hittade dock lite litteratur som specifikt behandlar denna möjliga koppling.
Ac:s roll har studerats i uppgifter som sannolikt kräver former av uppmärksamhet, t.ex. uppgifter som involverar visuell uppmärksamhet. Det är till exempel välkänt att acallosala populationer har studerats i samband med uppgifter som involverar visuell integration (Gazzaniga et al., 1962, 1965; Sperry, 1968; Levy et al., 1972). På senare tid försökte Corballis (1995) ta upp visuell integration och uppmärksamhet i ett försök att förklara tidigare diskrepanser i forskningen om acallosal. Corballis föreslog ett tvådelat visuellt uppmärksamhetssystem; en del (som involverar form) är automatisk och lokaliserad till varje hemisfär, och den andra (som involverar rörelse och plats) är frivillig och subkortikal. AAA-modellen kan bygga på denna observation och åtgärda de diskrepanser som observerats i litteraturen. Baserat på vår föreslagna modell finns det över flera nivåer av uppmärksamhet andra vägar än CC som kan förmedla informationsöverföring mellan hemisfärerna. En möjlighet är att involvera basala gangliakretsar som vi föreslår är potentiellt kritiska för att förmedla delning av uppmärksamhet mellan hemisfärerna (Franz, 2012). Genom att korsa CC försvinner i stort sett den direkta överföringen av uppmärksamhet via kortikala vägar (om inte AC förstås spelar en reservroll). Den uppmärksamhetsbaserade integrationen av visuell information kan dock fortfarande förmedlas via subkortikala vägar och/eller AC. På detta sätt skulle det tvådelade uppmärksamhetssystem som observerades av Corballis (1995) kunna vara närvarande på grund av den uppmärksamhetsbaserade förmedlingen av visuell integration via AC (och/eller basala ganglier och involverade kretsar: Franz, 2012).
Studier som involverar syn och uppmärksamhet behöver inte betraktas helt separat från det system som reglerar motoriska handlingar. Baserat på våra egna resultat (Franz, 2004; Franz och Packman, 2004) är olika former av uppmärksamhet (visuell och intern-nonvisuell) nära kopplade till motorisk handling, och det står klart att en möjlig nyckel till att förstå en potentiell uppmärksamhetsroll för AC ligger i kopplingen mellan handling, syn och uppmärksamhet. I en tidigare studie lät Franz (2004) normala deltagare rita cirklar bimanuellt under olika manipulationer av visuell återkoppling eller intern uppmärksamhet på varje hand. Storleken på de cirklar som ritades av en hand som fick en form av intern uppmärksamhet blev större på ett sätt som liknade effekter som uppstod vid närvaro av visuell återkoppling av den handen (i förhållande till ingen uppmärksamhet/visuell återkoppling). Vår AAA-modell innebär att uppmärksamhet är en nyckelkomponent i regleringen av den motoriska produktionen (liksom valet av vilken motorisk handling som planeras), och i den mån det finns bevis kan vi inte utesluta att AC spelar en roll i denna funktion. Ett direkt test av AC:s roll i någon kognitiv uppgift är dock inte trivialt.
Test av om AC deltar i kognitiv funktion är nästan omöjligt i neuropsykologisk forskning med tanke på att deltagare med lokaliserad AC inte är lätta att hitta (och vi känner inte till någon). Det kan därför vara mycket mer intuitivt att bedöma neurologiska skillnader i den normala AC och korrelera neurologiska data med beteendedata. Den presenterade traktografimetoden (som belyser AC) kan relativt enkelt tillämpas på stora urval av normalbefolkningen och kan helst kombineras med prestationer (baserade på uppgifter som utförs under separata testtillfällen från DTI-samlingen) hos samma individer med hjälp av specialdesignade kognitiva uppgifter. Här visar vi att DTI kan utföras och ge användbara resultat på ett litet urval av personer från den neurologiskt normala befolkningen. Det allmänna förslaget skulle vara att utföra liknande metoder på ett större urval och (som föreslaget ovan) korrelera resultaten (av ett proxymått för t.ex. AC:s täthet) med variabler på en kognitiv uppgift.
I figur 1 visar vi resultat av DTI-spårning av data från 10 kvinnliga deltagare som erhållits från ett offentligt DTI-dataset vid Johns Hopkins University (se tack för fullständig källa i Acknowledgments). Medelåldern var 22 år (SD = 2,5). Bilderna bearbetades med hjälp av FSL Diffusion Toolbox (Smith et al., 2004; Woolrich et al., 2009; Jenkinson et al., 2012) och förfarandena använde de som använts i tidigare studier av andra fiberbanor (FDT; Behrens et al., 2003a,b, 2007; Johansen-Berg et al., 2004). I korthet sammanfogades först bilderna till 4d-volymer för varje deltagare och skallarna strippades med hjälp av FSL:s Brain Extraction Tool (BET; Smith, 2002; Jenkinson et al., 2005). De extraherade bilderna korrigerades för mindre huvudrörelser och virvelströmmar som genererades av förändringar i fältets riktning. Diffusionstensorer anpassades för att generera bilder som visar primära diffusionsriktningar och FA-nivåer vid varje voxel som ska användas för färgkartor under maskering. Bayesiansk uppskattning av diffusionsparametrar som erhållits med hjälp av provtagningstekniker med korsande fibrer (BEDPOSTX; ”X” innebär korsande fibrer) genomfördes för att generera en fördelning av diffusionsstorlek och -riktning som tar hänsyn till två korsande fibrer för att senare genomföra traktografi över diffusionsvoxlar.
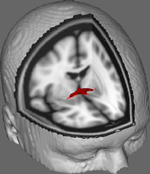
Figur 1. Genomsnittliga spårningsresultat över 10 kontrolldeltagare överlagrade på en standard T1-viktad MRI-bild med 2 mm isotropa voxlar för anatomisk referens.
Medelade spårningsresultat över alla 10 kontrolldeltagare överlagrades sedan på en standard T1-viktad MRI-bild med 2 mm isotropa voxlar för anatomisk referens. Gruppens genomsnittliga FA-förhållande var 0,3146 (SD = 0,0472). Dessa traktografiresultat avslöjar, vilket är viktigt, att strukturen hos AC kan belysas, och har därför potential för ytterligare studier som korrelerar resultatpoäng på beteendeuppgifter med FA-värden med hjälp av en större provstorlek.
Sammanfattningsvis finns det för närvarande sparsam litteratur, och av den tillgängliga litteraturen är det ingen som specifikt kopplar AC till processer som är relaterade till AAA-modellen, som är i fokus i vår tidigare målartikel (Franz, 2012). Med hjälp av ett förfarande som liknar det som föreslås här är det möjligt att testa större provstorlekar av normalbefolkningen och korrelera FA-värdena i DTI-resultaten (t.ex.) med en specifik beroende variabel som erhålls från en ändamålsenligt utvecklad uppgift om uppmärksamhet på handling (förfaranden för detta är en pågående utveckling i vårt laboratorium).
Intressekonfliktförklaring
Författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller ekonomiska relationer som skulle kunna tolkas som en potentiell intressekonflikt.
Acknowledgments
Vi är tacksamma för att Human Brain Project och National Research Resource Center (USA) och John Hopkins University har tillåtit offentlig användning av deras DTI-data. Vi är också tacksamma för generöst bidrag från Marsden Fund of the Royal Society of New Zealand (till Elizabeth A. Franz).
Corballis, M. (1995). Visuell integration i den delade hjärnan. Neuropsychologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). Tilldelning av uppmärksamhet vid inlärning av målstyrda handlingar: en kognitiv neurovetenskaplig ram med fokus på basala ganglier. Front. Psychol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., and Jones, E. (1971). Interhemisfäriska vägar vid avsaknad av corpus callosum. En experimentell studie av kommissurala förbindelser hos palshunden. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
Lämna ett svar