Bakteriecellernas struktur och funktion
On oktober 10, 2021 by adminBakteriecellernas struktur och funktion (sida 5)
(Detta kapitel har 10 sidor)
© Kenneth Todar, PhD
Cellvägg
Bakteriernas cellväggar förtjänar särskild uppmärksamhet av flera skäl:
1. De är en viktig struktur för livsduglighet, vilket beskrivs ovan.
2. De består av unika komponenter som inte finns någon annanstans i naturen.
3. De är en av de viktigaste platserna för angrepp av antibiotika.
4. De tillhandahåller ligander för vidhäftning och receptorplatser för läkemedel eller virus.
5. De orsakar sjukdomssymptom hos djur.
6. De möjliggör immunologisk distinktion och immunologisk variation mellan bakteriestammar.
De flesta prokaryoter har en hård cellvägg.Cellväggen är en väsentlig struktur som skyddar cellprotoplasten från mekanisk skada och från osmotisk bristning eller lysis.Prokaryoter lever vanligtvis i relativt utspädda miljöer så att ackumulationen av lösningsmedel i den prokaryotiska cellens cytoplasma vida överstiger den totala absoluta koncentrationen i den yttre miljön. Det osmotiska trycket mot plasmamembranets insida kan således motsvara 10-25 atm. Eftersom membranet är en känslig, plastisk struktur måste det hållas tillbaka av en yttervägg av poröst, styvt material med hög draghållfasthet. Ett sådant material är murein, den allestädes närvarande komponenten i bakteriella cellväggar.
Murein är en unik typ av petidoglykan, en polymer av disackarider (glykan) som är tvärbunden med korta kedjor av aminosyror (peptid). Det finns många typer av peptidoglykan. Alla bakteriella peptidoglykaner innehåller N-acetylmuraminsyra, som är den definierande beståndsdelen i murein. Archaea:s cellväggar kan bestå av protein, polysackarider eller peptidoglykanliknande molekyler, men de innehåller aldrig murein. Denna egenskap skiljer bakterier från arkéer.
I de Gram-positiva bakterierna (de som behåller det lila kristallvioletta färgämnet när de utsätts för Gram-färgningsförfarandet) består cellväggen av flera lager peptidoglykan. Liktydigt med peptidoglykanskikten löper en grupp molekyler som kallas teikosyror och som är unika för den grampositiva cellväggen (figur 14). 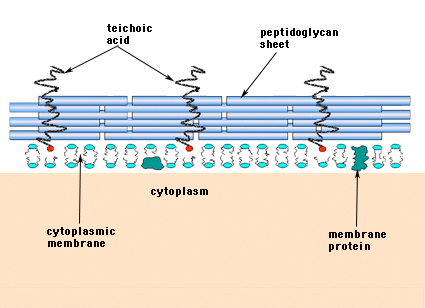
Figur 14. Strukturen hos den grampositiva bakteriens cellvägg. Väggen är relativt tjock och består av många lager peptidoglykan varvat med teikosyror som löper vinkelrätt mot peptidoglykanskikten.
I gramnegativa bakterier (som inte behåller kristallviolett) består cellväggen av ett enda lager peptidoglykaner som omges av en membranstruktur som kallas det yttre membranet.Det yttre membranet hos gramnegativa bakterier innehåller alltid en unik komponent, lipopolysackarid (LPS eller endotoxin), som är giftig för djur. Hos gramnegativa bakterier brukar det yttre membranet betraktas som en del av cellväggen (figur 15).

Figur 15. Strukturen hos den gramnegativa cellväggen. Väggen är relativt tunn och innehåller mycket mindre peptidoglykan än den Gram-positiva väggen. även teikosyror saknas. Den gramnegativa cellväggen består dock av ett yttre membran som ligger utanför peptidoglykanlagret. Det yttre membranet är fäst vid peptidoglykanskiktet genom en unik grupp av lipoproteinmolekyler.
I de grampositiva bakterierna är cellväggen tjock (15-80 nanometer) och består av flera lager peptidoglykan. I gramnegativa bakterier är cellväggen relativt tunn (10 nanometer) och består av ett enda lager peptidoglykaner som omges av ett yttre membran.
Peptidoglykanstrukturen och -arrangemanget i E.coli är representativt för alla Enterobacteriaceae, liksom för många andra gramnegativa bakterier. Glykanryggen består av alternerande molekyler av N-acetylglukosamin (G) och N-acetylmuraminsyra (M) som är sammankopplade genom en beta 1,4-glykosidbindning. N-acetylmuraminsyras (M) 3-kolämne är substituerat med en laktyletergrupp som härrör från pyruvat. Laktyletern förbinder glykanryggen med en peptidsidokedja som innehåller L-alanin (L-ala), D-glutamat (D-glu), diaminopimelsyra (DAP) och D-alanin (D-ala). MurNAc är unikt för bakteriella cellväggar, liksom D-glu, DAP och D-ala. Muraminsyraunderenheten i E. coli visas i figur 16 nedan.

Figur 16. Strukturen av muraminsyraunderenheten i peptidoglykanen hos Escherichia coli.Detta är den typ av murein som finns i de flesta gramnegativa bakterier. Glykanryggen är en upprepad polymer av två aminosocker, N-acetylglukosamin (G) och N-acetylmuraminsyra (M). Till N-acetylmuraminsyran är enetrapeptid som består av L-ala-D-glu-DAP-D-ala fäst vid N-acetylmuraminsyran. b. Förkortad struktur för muraminsyraunderenheten. c. Närliggande tetrapeptidsidekedjor kan vara kopplade till varandra genom en interpeptidbindning mellan DAP på den ena kedjan och D-ala på den andra. d. Molekylens polymera form.
Mureinsträngar byggs upp i periplasman av cirka 10 muraminsyraunderenheter. När deras närhet tillåter det kan de tetrapeptidkedjor som sticker ut från glykanryggen tvärbindas genom en interpeptidbindning mellan en fri aminogrupp på DAP och en fri karboxygrupp på en närliggande D-ala.Peptidoglykanen samlas på utsidan av plasmamembranet med hjälp av en grupp periplasmatiska enzymer som är transglykosylaser, transpeptidaser och karboxypeptidaser. Penicillin och besläktade beta-laktamantibiotika verkar genom att blockera transpeptidas- och karboxypeptidasenzymerna under deras uppbyggnad av mureincellväggen. Därför sägs betalakatamantibiotika ”blockera cellväggssyntesen” i bakterierna.
Peptidoglykanmolekylens glykanryggrad kan klyvas av ett enzym som kallas lysozym och som finns i djurserum, vävnader och sekret samt i den fagocytiska lysosomen. Lysozymets funktion är att lysera bakterieceller som ett konstitutivt försvar mot bakteriella patogener. Vissa grampositiva bakterier är mycket känsliga för lysozym och enzymet är ganska aktivt vid låga koncentrationer.Lachrymmarker (tårar) kan spädas ut 1:40 000 och behåller förmågan att lysera vissa bakterieceller. Gramnegativa bakterier är mindre känsliga för lysozym eftersom deras peptidoglykan är skyddad av ytterhinnan. Den exakta platsen för lysozymets klyvning är beta 1,4-bindningen mellan N-acetylmuraminsyra (M) och N-acetylglukosamin (G), så att den underenhet av duraminsyra som visas i figur 16(a) är resultatet av lysozymets verkan på bakteriell peptidoglykan.
Inom Gram-positiva bakterier finns det ett stort antal olikapeptidarrangemang bland peptidoglykanerna. Den bäst studerade är denurein hos Staphylococcus aureus som visas i figur 17 nedan. I stället för DAP (i E. coli) finns diaminosyran L-lysin (L-lys), och i stället för interpeptidbindningen (i Gram-negativa) finns en interpeptidbro av aminosyror som förbinder en fri aminogrupp på lysin med en fri karboxygrupp på D-ala i en närliggande tetrapeptidsidokedja. Detta arrangemang möjliggör tydligen en mer frekvent korsbindning mellan närliggande tetrapeptidsidokedjor. I S. aureus är interpeptidbryggan en peptiddekoration bestående av fem glycinmolekyler (kallad pentaglycinbrygga). Samlingen av interpeptidbryggan i Gram-positivt murein hämmas av beta-laktamantibiotika på samma sätt som interpeptidbindningen i Gram-negativt murein. Grampositiva bakterier är känsligare för penicillin än gramnegativa bakterier eftersom peptidoglykanen inte skyddas av ett yttre membran och är en rikligare molekyl. I grampositiva bakterier kan peptidoglykanerna variera i fråga om vilken aminosyra som ersätter DAP eller L-lys i position 3 i tetrapeptiden och i fråga om den exakta sammansättningen av interpeptidbryggan. Minst åtta olika typer av peptidoglykaner finns i Gram-positiva bakterier.
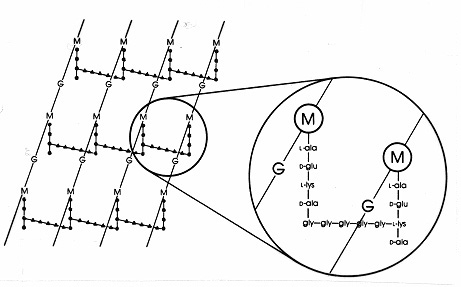
Figur 17. Schematisk bild av peptidoglykanen hos Staphylococcus aureus. G = N-acetyl-glukosamin, M = N-acetyl-muraminsyra, L-ala = L-alanin, D-ala = D-alanin, D-glu = D-glutaminsyra, L-lys = L-lysin. Detta är en typ av murein som finns i grampositiva bakterier. Jämfört med E. coli-peptidoglykanen (figur 7) finns L-lys i stället för DAP (diaminopimelinsyra) i tetrapeptiden.L-lys’ fria aminogrupp är ersatt med en glycinpentapeptid (gly-gly-gly-gly-gly-gly-gly-) som sedan blir en interpeptidbrygga som bildar en länk med en karboxygrupp från D-ala i en intilliggande tetrapeptidsidokedja. Grampositiva peptidoglykaner skiljer sig från art till art, främst med avseende på aminosyrorna i den tredje positionen i tetrapeptidkedjan och aminosyrasammansättningen i interpeptidbryggan.
Gramnegativa bakterier kan innehålla ett enda molekylärt lager av murein i sina cellväggar, medan grampositiva bakterier anses ha flera lager eller ”omslag” av peptidoglykan. I nära anslutning till lagren av peptidoglykan i grampositiva bakterier finns en grupp molekyler som kallas teikosyror. Teikosyror är linjära polymerer av polyglycerol eller polyribitol som är substituerade med fosfater och ett fåtal aminosyror och sockerarter. Teikosyrepolymererna är ibland förankrade i plasmamembranet (lipoteikosyra, LTA) och verkar vara riktade utåt i rät vinkel mot peptidoglykanlagren. Funktionerna hos teichonsyra är inte kända. De är nödvändiga för att grampositiva bakterier skall vara livskraftiga i det vilda. En idé är att de utgör en kanal med regelbundet orienterade negativa laddningar för att föra in positivt laddade ämnen i det komplicerade peptidoglykan-nätverket. En annan teori är att teikosyrorna på något sätt är inblandade i regleringen och sammansättningen av muraminsyraunderenheter på utsidan av plasmamembranet. Det finns exempel, särskilt hos streptokocker, där teikosyror har varit inblandade i bakteriernas vidhäftning till vävnadsytor.
kapitlet fortsätter
förra sidan
.
Lämna ett svar