Översta olivariekärnan
On november 12, 2021 by admin| Hjärnan: Superior olivary complex | ||
|---|---|---|
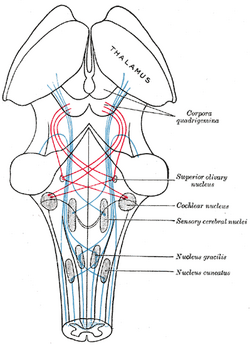 |
||
| Schema som visar förloppet av fibrerna i lemniscus; medial lemniscus i blått, lateralt i rött. (Superior olivary nucleus är märkt i mitten till höger.) | ||
| ] | ||
| Latin | nucleus olivaris superior | |
| Grays | subjekt #187 787 | |
| del av | ||
| Komponenter | ||
| Arterie | ||
| Vein | ||
| BrainInfo/UW | {{{{BrainInfoType}}}-565 | |
| MeSH | ||
Det övre olivariekomplexet (eller SOC eller superior olive) är en liten massa grå substans som ligger på dorsalytan av den laterala delen av trapezoidkroppen. Det är litet hos människor, men välutvecklat hos vissa djur, och uppvisar liknande struktur som den nedre olivariekärnan, och ligger omedelbart ovanför den. Vissa av fibrerna i trapezoidkroppen slutar genom att arborisera runt cellerna i denna kärna, medan andra uppstår från dessa celler.
Fysiologi
Den överlägsna olivariekärnan spelar ett antal roller i hörseln. Medial superior olive (MSO) är en specialiserad kärna som tros mäta tidsskillnaden för ljudets ankomst mellan öronen (den interaurala tidsskillnaden eller ITD). ITD är en viktig ledtråd för att bestämma azimut för lågfrekventa ljud, dvs. att lokalisera dem på det azimutala planet – deras grad till vänster eller höger.
Den laterala superior oliv (LSO) tros vara involverad i mätningen av ljudets nivåskillnad mellan öronen (den interaurala nivåskillnaden eller ILD). ILD är en andra viktig ledtråd när det gäller att bestämma azimut av högfrekventa ljud.
Förhållande till hörselsystemet
Det överordnade olivkomplexet är i allmänhet beläget i pons, men hos människan sträcker sig från den rostrala märgen till mitten av pons och tar emot projektioner främst från den anteroventrala cochlearkärnan via den ventrala akustiska strimman, även om den posterventrala kärnan också projicerar dit via den intermediära akustiska strimman. Det är på denna plats som de första stora binaurala interaktionerna sker.
Primära kärnor
Det övre olivariekomplexet är uppdelat i tre primära kärnor, MSO, LSO och den mediala kärnan i trapezoidkroppen, och flera mindre periolivära kärnor. Dessa tre kärnor är de mest studerade och därför bäst förstådda. Typiskt sett anses de bilda den stigande azimutala lokaliseringsvägen.
Medial superior olive (MSO)
Den mediala superiora olivkärnan tros hjälpa till att lokalisera azimuten av ett ljud, det vill säga vinkeln till vänster eller höger där ljudkällan är belägen. Ens första instinkt kan vara att tro att denna kärna omfattar vertikal information, men så är inte fallet; den information som bearbetas i MSO är begränsad till det horisontella planet. De fusiforma cellerna projicerar inte till något i nivå med pons och kommer först in i bilden vid inferior colliculus. Endast horisontell information förekommer, men den kommer från två olika öronkällor, vilket underlättar lokaliseringen av ljudet på azimutaxeln. Det sätt på vilket den övre olivoljan gör detta är genom att mäta tidsskillnaderna mellan två öronsignaler som registrerar samma stimulus. Att färdas runt huvudet tar ungefär 700 μs, och man antar att den mediala superior olive kan registrera detta. I själva verket har man observerat att människor kan upptäcka interaurala skillnader ner till 10 mikrosekunder. Kärnan är tonotopiskt organiserad, men den azimutala receptiva fältprojektionen är ”sannolikt en komplex, icke-linjär karta”.
Projektionerna från den mediala övre oliven avslutas tätt i den ipsilaterala centrala kärnan i inferior colliculus. Majoriteten av dessa axoner anses vara ”rundformade” eller typ R. Dessa R-axoner är mestadels glutamatergiska och innehåller runda synaptiska vesiklar och bildar asymmetriska synaptiska förbindningar.
- Detta är den största av kärnorna och innehåller hos människan cirka 15 500 neuroner .
- Varje MSO tar emot lågfrekventa bilaterala ingångar från höger och vänster AVCNs.
- Utgången går till den ipsilaterala laterala lemniscus och i slutändan till inferior colliculus.
- Den MSO reagerar bättre på binaurala stimuli.
- Dess huvudsakliga funktion innebär upptäckt av skillnader i ankomsttid för ljud till de två öronen, vilket är en del av lokaliseringsprocessen (ITD).
Lateral superior olive (LSO)
Denna oliv har liknande funktioner som medial superior olive, men använder intensitet för att lokalisera ljudkällan. Detta är den del av hjärnstammen som markerar att det starkare ljudet från vänster öra ligger på vänster sida. Den laterala oliven får input från båda cochlearkärnorna, från ipsilaterala AVCN sfäriska bushy-celler och kontralaterala AVCN globulära bushy-celler via den ipsilaterala MNTB. MNTB:s huvudceller är glycinergiska, vilket ger en källa till hämning till LSO och utgör grunden för ILD-känsligheten. Projektioner från båda cochlearkärnorna är huvudsakligen högfrekventa, och dessa frekvenser representeras sedan av majoriteten av LSO-neuronerna (>2/3 över 2-3 kHz hos katt). Ytterligare ingångar kommer från den ipsilaterala LNTB (glycinergiskt, se nedan), som ger hämmande information från den ipsilaterala cochlearkärnan. En annan eventuellt hämmande input kommer från ipsilaterala icke-sfäriska celler i AVCN. Dessa celler är antingen globulära buskiga eller multipolära (stellate). Någon av dessa två ingångar kan utgöra grunden för ipsilateral hämning som ses i responskartor som flankerar den primära excitationen, vilket skärper enhetens frekvensavstämning.
LSO projicerar bilateralt till den centrala kärnan i inferior colliculus (ICC). Ipsilaterala projektioner är främst hämmande (glycinergiska) och kontralaterala excitatoriska. Ytterligare projektionsmål omfattar de dorsala och ventrala kärnorna i laterala lemniscus (DNLL & VNLL). De GABAergiska projektionerna från DNLL utgör den primära källan till GABA i den auditiva hjärnstammen och projicerar bilateralt till ICC och till den kontralaterala DNLL. Dessa konvergerande excitatoriska och hämmande förbindelser kan verka för att minska nivåberoendet av ILD-känsligheten i ICC jämfört med LSO.
Utvidare projektioner bildar Lateral Olivocochlear Bundle (LOC), som innerverar cochleära inre hårceller. Dessa projektioner tros ha en lång tidskonstant och fungerar för att normalisera ljudnivån som detekteras av varje öra för att underlätta ljudlokalisering. Det finns avsevärda artskillnader: LOC-projektionsneuronerna är fördelade inom LSO hos gnagare och omger LSO hos rovdjur (t.ex. katt).
Medial Nucleus of Trapezoid Body (MNTB)
- MNTB är den minsta delen av SOC; närvaron av MNTB i den mänskliga hjärnstammen är tveksam.
- Varje neuron får endast högfrekventa inflöden via ett Calyx of held från den kontralaterala AVCN, och dess utgång projicerar till den ipsilaterala LSO.
- Det finns två responstyper som hittas: en ”choppertyp” som liknar spindelceller i AVCN och en primär typ som liknar de Bushy Cells i AVCN.
Periolivary Nuclei
SOC består av mellan sex och nio periolivary nuclei, beroende på vilken forskare som åberopas, och de benämns vanligen baserat på deras placering i förhållande till de primära kärnorna. Dessa kärnor omger var och en av de primära kärnorna och bidrar till både det uppåtgående och det nedåtgående hörselsystemet. Dessa kärnor utgör också källan till olivocochlearbunten, som innerverar snäckan. Hos marsvinet är de uppåtgående projektionerna till inferior colliculi huvudsakligen ipsilaterala (>80 %), och den största enskilda källan kommer från SPON. Även de ventrala kärnorna (RPO, VMPO, AVPO, & VNTB) är nästan helt ipsilaterala, medan de övriga kärnorna projicerar bilateralt.
| Namn | Katt | Guinea Pig | Rat | Mus | ||
|---|---|---|---|---|---|---|
| LSO | X | X | X | X | X | |
| MSO | X | X | X | X | ||
| MNTB | X | X | X | X | X | X |
| LNTB | X | X | ”LVPO” | X | ||
| ALPO | X | X | ||||
| PVPO | X | X | ||||
| PPO | X | X | ”CPO” | |||
| VVLPO | X | |||||
| DPO | X | X | X | |||
| DLPO | X | X | X | |||
| VTB | X | X | ”MVPO” | X | ||
| AVPO | X | |||||
| VMPO | X | X | ||||
| RPO | X | X | ||||
| SPN | ”DMPO” | X | X | X | X |
Ventral Nucleus of Trapezoid Body (VNTB)
- VENTRAL Nucleus of Trapezoid Body (VNTB) är en liten kärna som ligger lateralt till MNTB, och ventral till MSO.
- Denna kärna består av en heterogen cellpopulation och projicerar till många hörselkärnor och bildar det mediala olivocochleära buntet (MOC) som innerverar cochleära yttre hårceller . Dessa celler innehåller elektromotila fibrer och fungerar som mekaniska förstärkare/avdämpare i cochlea.
- Kärnan projicerar till båda IC, utan att några celler projicerar bilateralt.
Lateral Nucleus of the Trapezoid Body (LNTB)
- Lokaliserad ventralt till LSO
- AVCN sfäriska buskiga celler projicerar kollateraler bilateralt, och globulära buskiga celler projicerar kollateraler ipsilateralt till LNTB-neuroner.
- Cellerna är immunoreaktiva för glycin och är retrogrademärkta efter injektion av tritiat glycin i LSO
- Kärnan projicerar till både IC, med få celler som projicerar bilateralt, samt till den ipsilaterala LSO.
- Stora multipolära celler projicerar till cochlearkärnan, men inte till IC, hos både katt och marsvin.
- Inputs sker ofta via slutbulber av held, vilket ger mycket snabb signaltransduktion.
Superior Periolivary Nucleus (SPON) (Dorsomedial Periolivary Nucleus (DMPO))
- Lokaliserad direkt dorsalt i förhållande till MNTB
- Hos råtta är SPON en homogen GABAerg kärna. Dessa tonotopiskt organiserade neuroner får excitatoriska ingångar från octopus och multipolära celler i den kontralaterala ventrala cochlearkärnan, en glycinerg (hämmande) ingång från den ipsilaterala MNTB, en okänd GABAerg (hämmande) ingång, och projicerar till den ipsilaterala ICC. De flesta neuronerna reagerar endast vid offset av ett stimulus, kan faslåsas till AM-stimuli upp till 200 Hz och kan ligga till grund för ICC:s varaktighetsselektivitet. SPON-neuronerna tar inte emot nedåtgående ingångar från IC och projicerar inte till cochlea eller cochlear nucleus, vilket många periolivära kärnor gör.
- I marsvin projicerar runda till ovala multipolära celler till båda IC, med många celler som projicerar bilateralt. De mer långsträckta cellerna som projicerar till cochlearkärnan projicerar inte till ICC. Det verkar finnas två populationer av celler, en som projicerar ipsilateralt och en som projicerar bilateralt.
- Majoriteten av informationen hade kommit från gnagare SPON, på grund av kärnans framträdande storlek hos dessa arter, med mycket få studier har gjorts på DMPO hos katt, och ingen av dem var omfattande.
Dorsal Periolivary Nucleus (DPO)
- Lokaliserad dorsalt och medial till LSO
- Innehåller både EE (exciteras av båda öronen) och E0 (exciteras endast av det kontralaterala örat) enheter.
- Neuronerna är tonotopiskt organiserade och högfrekventa
- Kan tillhöra en enda kärna tillsammans med DLPO
- Kärnan projicerar till båda IC, utan celler som projicerar bilateralt.
Dorsolateral Periolivary Nucleus (DLPO)
- Lokaliserad dorsalt och lateralt till LSO
- Innehåller både EE (exciteras av båda öronen) och E0 (exciteras endast av det kontralaterala örat) enheter.
- Neuronerna är tonotopiskt organiserade och lågfrekventa.
- Kan tillhöra en enda kärna tillsammans med DPO
- Kärnan projicerar till båda IC, med få celler som projicerar bilateralt.
Ventrolateral Periolivary Nucleus (VLPO)
- Lokaliserad ventralt till och inom LSO:s ventrala hillus
- Innehåller både EI (exciteras av kontralaterala och hämmas av ipsilaterala örat) och E0 (exciteras endast av det kontralaterala örat) enheter.
- Neuronerna är tonotopiskt organiserade och högfrekventa.
Anterolateral Periolivary Nucleus (ALPO)
- Kärnan projicerar till båda IC, utan celler som projicerar bilateralt.
- Stora multipolära celler projicerar till cochlearkärnan, men inte till IC, hos både katt och marsvin.
Ventromedial Periolivary Nucleus (VMPO)
- Lokaliserad mellan MSO och MNTB.
- Sänder projektioner till ICC bilateralt.
- Kärnan projicerar till båda IC, utan att några celler projicerar bilateralt.
Rostral Periolivary Nucleus (RPO) (Anterior Periolivary Nucleus (APO))
- Lokaliserad mellan den rostrala polen av MSO och VNLL
Caudal Periolivary Nucleus (CPO) (Posterior Periolivary Nucleus (PPO))
- Lokaliserad mellan den kaudala polen av MSO och ansiktskärnan (7N)
Posteroventral Periolivary Nucleus (PVPO)
- Kärnan projicerar till båda IC, utan celler som projicerar bilateralt.
Patofysiologi
Vid obduktion av en 21-årig kvinna med autism, epilepsi och mental retardation fann man en nästan fullständig frånvaro av den övre oliven.
Se även
- Olivkroppen
- Kulesza RJ, Cytoarchitecture of the human superior olivary complex: Medial and lateral superior olive. Hearing Research 225(2007) 80-90
- 2,0 2,1 Oliver DL, et al. Axonala projektioner från den laterala och mediala överordnade oliven till inferior colliculus hos katten: en studie med hjälp av elektronmikroskopisk autoradiografi. J Comp Neurol. 1995 Sep 11;360(1):17-32 Citerar fel: Invalid
<ref>tag; name ”R4” defined multiple times with different content - Cajal, S. R. Y. and L. Azoulay (1909). Histologie du système nerveux de l’homme et des vertébrés. Paris, Maloine.
- 4.0 4.1 Kandel, et al Principles of Neuroscience. Fourth ed. pp 591-624. Copyright 2000, av McGraw-Hill Co. Citatfel: Felaktig
<ref>-tagg; namnet ”R2” definierat flera gånger med olika innehåll - Kulesza, R. J., Jr. (2007). ”Cytoarkitektur hos människans övre olivariekomplex: medial och lateral övre oliv”. Hear Res 225(1-2): 80-90.
- Tsuchitani, C. och J. C. Boudreau (1967). ”Kodning av stimulusfrekvens och intensitet av S-segmentcellerna i kattens övre oliv.” J Acoust Soc Am 42(4): 794-805.
- 8.0 8.1 8.2 Citatfel: Invalid
<ref>tag; name ”R10” defined multiple times with different content - Wu, S. H. and J. B. Kelly (1994). ”Fysiologiska bevis för ipsilateral hämning i lateral superior olive: synaptiska svar i hjärnskiva av mus”. Hear Res 73(1): 57-64.
- Brownell, W. E., P. B. Manis, et al. (1979). ”Ipsilaterala hämmande reaktioner i kattens laterala övre oliv.” Brain Res 177(1): 189-93.
- Darrow, K. N., S. F. Maison, et al. (2006). ”Cochlear efferent feedback balanserar interaural känslighet”. Nat Neurosci 9(12): 1474-6.
- Warr, W. B. och J. J. Guinan, Jr. (1979). ”Efferent innervation av cortiorganet: två separata system”. Brain Res 173(1): 152-5.
- 13.0 13.1 Schofield, B. R. och N. B. Cant (1991). ”Organisering av det övre olivariekomplexet hos marsvinet. I. Cytoarkitektur, histokemi för cytokromoxidas och dendritisk morfologi”. J Comp Neurol 314(4): 645-70. Citatfel: Invalid
<ref>tag; name ”R7” defined multiple times with different content - 14.0 14.1 14.2 14.3 14.4 14.5 14.6 14.7 14.8 Illing, R. B., K. S. Kraus, et al. ”Plasticity of the superior olivary complex”. Microsc Res Tech 51(4): 364-81. Citatfel: Fel: Felaktig
<ref>-tagg; namnet ”R5” har definierats flera gånger med olika innehåll: Felaktig<ref>-tagg; namnet ”R5” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Invalid<ref>tag: Felaktig<ref>-tagg; namnet ”R5” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Felaktig<ref>-tagg; namnet ”R5” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Felaktig<ref>-tagg; namnet ”R5” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Felaktig<ref>-tagg; namnet ”R5” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Invalid<ref>tag; name ”R5” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R5” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R5” defined multiple times with different content - Warr, W. B. and J. E. Beck (1996). ”Multipla projektioner från den ventrala kärnan i trapetskroppen hos råttan”. Hear Res 93(1-2): 83-101.
- 16.00 16.01 16.02 16.03 16.04 16.05 16.06 16.06 16.07 16.08 16.09 16.10 Schofield, B. R. och N. B. Cant (1992). ”Organisering av det övre olivariekomplexet hos marsvinet: II. Projektionsmönster från de periolivära kärnorna till inferior colliculus”. J Comp Neurol 317(4): 438-55. Cite error: Fel i
<ref>taggen; namnet ”R6” definierat flera gånger med olika innehåll Cite error: Invalid<ref>tagg: Felaktig<ref>-tagg; namnet ”R6” har definierats flera gånger med olika innehåll: Felaktig<ref>-tagg; namnet ”R6” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Felaktig<ref>-tagg; namnet ”R6” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Felaktig<ref>-tagg; namnet ”R6” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Felaktig<ref>-tagg; namnet ”R6” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Felaktig<ref>-tagg; namnet ”R6” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Felaktig<ref>-tagg; namnet ”R6” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Felaktig<ref>-tagg; namnet ”R6” definierat flera gånger med olika innehåll Cite error: Felaktig<ref>-tagg: Invalid<ref>tag; name ”R6” defined multiple times with different content - Smith, P. H., P. X. Joris, et al. (1993). ”Projektioner av fysiologiskt karakteriserade sfäriska bushy cell axoner från cochlearkärnan hos katten: bevis för fördröjningslinjer till den mediala superiöra oliven”. J Comp Neurol 331(2): 245-60.
- Wenthold, R. J., D. Huie, et al. (1987). ”Glycin immunoreaktivitet lokaliserad i cochlearkärnan och det övre olivariekomplexet”. Neuroscience 22(3): 897-912.
- 19.0 19.1 Adams, J. C. (1983). ”Cytologi av periolivära celler och organisationen av deras projektioner hos katten”. J Comp Neurol 215(3): 275-89. Cite error: Invalid
<ref>tag; name ”R8” defined multiple times with different content - Friauf, E. and J. Ostwald (1988). ”Divergerande projektioner hos fysiologiskt karakteriserade neuroner i råttans ventrala cochlearkärna som visas genom intraaxonal injektion av pepparrotsperoxidas”. Exp Brain Res 73(2): 263-84.
- Kulesza, R. J., Jr. och A. S. Berrebi (2000). ”Superior paraolivary nucleus of the rat is a GABAergic nucleus”. J Assoc Res Otolaryngol 1(4): 255-69.
- Kulesza, R. J., Jr., G. A. Spirou, et al. (2003). ”Fysiologiska responsegenskaper hos neuronerna i den övre paraolivära kärnan hos råttan”. J Neurophysiol 89(4): 2299-312.
- WHITE JS, WARR WB. Det dubbla ursprunget till det olivocochleära buntet hos albino-råttan. J. Comp. Neurol. 219:203-214, 1983.
- Guinan, J. J., Jr., S. S. Guinan, et al. (1972). ”Enskilda auditiva enheter i det övre olivariekomplexet. I. Svar på ljud och klassificeringar baserade på fysiologiska egenskaper”. Int J Neurosci 4: 101-20.
- Tsuchitani, C. (1977). ”Funktionell organisation av laterala cellgrupper i kattens övre olivariekomplex”. J Neurophysiol 40(2): 296-318.
- Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996). Embryologiskt ursprung för autism: utvecklingsavvikelser i kranialnervens motoriska kärnor. J Comp Neurol 370 (2): 247-61.
- MedEd at Loyola Neuro/frames/nlBSs/nl27fr.htm
- Image at med.utah.edu (pick 9b. Pons)
Den här artikeln baserades ursprungligen på en post från en public domain edition av Gray’s Anatomy. Därför kan en del av informationen vara föråldrad. Redigera artikeln om så är fallet, och ta gärna bort detta meddelande när det inte längre är relevant.
|
v-d-e
Hjärna: rhombencephalon (bakhjärna) |
||||||
|---|---|---|---|---|---|---|
| Metencephalon/ pons |
|
Dorsal/
(tegmentum) |
Ventralt/
(bas) |
Raphe/reticular
|
||
| Myelencephalon/ medulla |
|
Dorsal
|
Ventral
|
Raphe/reticular
|
||
|
v-d-e
Sinnessystem: Hörsel- och vestibulära system (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Uttre örat |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, örsnibben) – Hörselgången – Öronmuskulatur Hörntrumma (Umbo, Pars flaccida) |
|||||||||||
| Mellanörat |
|
|||||||||||
| Innerörat/ (membranös labyrint, benig labyrint) |
|
|||||||||||
| {| class=”navbox collapsible nowraplinks” style=”margin:auto;” | ||||||||||||
|
—
|
||||||||||||
|
||||||||||||
|}
Denna sida använder sig av Creative Commons-licensierat innehåll från Wikipedia (visa författare).
Lämna ett svar