Teoria Hebbian
On decembrie 14, 2021 by admin Evaluare | Biopsihologie | Comparată |Cognitivă | Dezvoltare | Limbaj | Diferențe individuale | Personalitate | Filosofie | Socială |
Metode | Statistică | Clinică | Educațională | Industrială | Industrială | Articole profesionale | Psihologie mondială |
Psihologie cognitivă:Atenție – Luare de decizii -Învățare – Judecată -Memorie – Motivație – Percepție – Raționament – Gândire -Procese cognitiveCogniție -OutlineIndex
Teoria Hebiană descrie un mecanism de bază pentru plasticitatea sinaptică în care o creștere a eficacității sinaptice apare din stimularea repetată și persistentă a celulei postsinaptice de către celula presinaptică. Introdusă de Donald Hebb în 1949, este numită și regula lui Hebb, postulatul lui Hebb și teoria asamblării celulare și afirmă:
Să presupunem că persistența sau repetarea unei activități reverberante (sau „urmă”) tinde să inducă modificări celulare de durată care se adaugă la stabilitatea sa…. Atunci când un axon al celulei A este suficient de aproape pentru a excita o celulă B și participă în mod repetat sau persistent la aprinderea acesteia, un anumit proces de creștere sau o schimbare metabolică are loc în una sau în ambele celule, astfel încât eficiența lui A, ca una dintre celulele care o aprinde pe B, este crescută.
Teoria este adesea rezumată ca „celulele care trag împreună, se conectează împreună”, deși aceasta este o simplificare excesivă a sistemului nervos care nu trebuie luată în mod literal, precum și faptul că nu reprezintă cu exactitate afirmația originală a lui Hebb cu privire la schimbările de intensitate a conectivității celulelor. Teoria este evocată în mod obișnuit pentru a explica unele tipuri de învățare asociativă, în care activarea simultană a celulelor duce la creșteri pronunțate ale puterii sinaptice. O astfel de învățare este cunoscută sub numele de învățare Hebbian.
Engramele Hebbian și teoria asamblării celulare
Teoria Hebbian se referă la modul în care neuronii s-ar putea conecta între ei pentru a deveni engrame. Teoriile lui Hebb cu privire la forma și funcția ansamblurilor celulare pot fi înțelese din următoarele:
„Ideea generală este una veche, că orice două celule sau sisteme de celule care sunt active în mod repetat în același timp vor tinde să devină „asociate”, astfel încât activitatea uneia facilitează activitatea celeilalte.” (Hebb 1949, p. 70) „Când o celulă ajută în mod repetat la aprinderea alteia, axonul primei celule dezvoltă noduri sinaptice (sau le mărește, dacă există deja) în contact cu soma celei de-a doua celule.” (Hebb 1949, p. 63)
Gordon Allport postulează idei suplimentare cu privire la teoria asamblării celulare și rolul acesteia în formarea engramelor, pe linia conceptului de autoasociere, descris după cum urmează:
„Dacă intrările unui sistem determină apariția repetată a aceluiași model de activitate, setul de elemente active care constituie acel model va deveni din ce în ce mai puternic interasociat. Adică, fiecare element va avea tendința de a activa fiecare alt element și (cu ponderi negative) de a dezactiva elementele care nu fac parte din tipar. Altfel spus, modelul ca întreg va deveni „auto-asociat”. Putem numi un tipar învățat (auto-asociat) o engramă.” (Hebb 1949, p. 44)
Teoria Hebbian a fost baza principală pentru opinia convențională conform căreia, atunci când sunt analizate de la un nivel holistic, engramele sunt rețele neuronale sau rețele neuronale.
Lucrările din laboratorul lui Eric Kandel au furnizat dovezi pentru implicarea mecanismelor de învățare Hebbian la nivelul sinapselor la gasteropodul marin Aplysia californica.
Experimentele privind mecanismele Hebbian de modificare a sinapselor la nivelul sinapselor sistemului nervos central al vertebratelor sunt mult mai dificil de controlat decât experimentele cu sinapsele relativ simple ale sistemului nervos periferic studiate la nevertebratele marine. O mare parte din lucrările privind modificările sinaptice de lungă durată între neuronii vertebratelor (cum ar fi potențarea pe termen lung) implică utilizarea stimulării experimentale non-fiziologice a celulelor cerebrale. Cu toate acestea, unele dintre mecanismele de modificare a sinapselor relevante din punct de vedere fiziologic care au fost studiate în creierul vertebratelor par a fi exemple de procese hebiene. Un astfel de studiu trece în revistă rezultatele unor experimente care indică faptul că modificările de lungă durată ale intensităților sinaptice pot fi induse de o activitate sinaptică relevantă din punct de vedere fiziologic care funcționează atât prin mecanisme Hebbian, cât și non-Hebbian
Principii
Din punctul de vedere al neuronilor artificiali și al rețelelor neuronale artificiale, principiul lui Hebb poate fi descris ca o metodă de determinare a modului de modificare a ponderilor dintre neuronii model. Ponderea dintre doi neuroni crește dacă cei doi neuroni se activează simultan – și se reduce dacă se activează separat. Nodurile care tind să fie fie ambele pozitive sau ambele negative în același timp au ponderi pozitive puternice, în timp ce cele care tind să fie opuse au ponderi negative puternice.
Acest principiu original este poate cea mai simplă formă de selecție a ponderilor. În timp ce acest lucru înseamnă că poate fi codificat relativ ușor într-un program de calculator și utilizat pentru a actualiza ponderile unei rețele, el interzice, de asemenea, numărul de aplicații ale învățării Hebbian. În prezent, termenul de învățare Hebbian se referă, în general, la o anumită formă de abstractizare matematică a principiului original propus de Hebb. În acest sens, învățarea Hebbian implică ajustarea ponderilor dintre nodurile de învățare astfel încât fiecare greutate să reprezinte mai bine relația dintre noduri. Ca atare, multe metode de învățare pot fi considerate ca fiind într-o oarecare măsură de natură hebbiană.
Cele ce urmează reprezintă o descriere formulaică a învățării hebiene: (rețineți că sunt posibile multe alte descrieri)
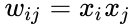
unde 
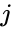
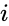
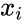
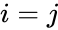
O altă descriere prin formulă este:
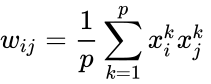
, unde 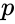

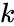
O variantă a învățării Hebbian care ia în considerare fenomene precum blocajul și multe alte fenomene de învățare neuronală este modelul matematic al lui Harry Klopf. Modelul lui Klopf reproduce foarte multe fenomene biologice și este, de asemenea, simplu de implementat.
Generalizare și stabilitate
Regula lui Hebb este adesea generalizată sub forma
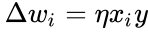
sau variația în 
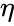
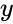
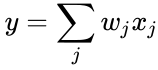
și simplificarea din secțiunea anterioară ia atât rata de învățare cât și ponderile de intrare ca fiind 1. Această versiune a regulii este în mod clar instabilă, deoarece în orice rețea cu un semnal dominant, greutățile sinaptice vor crește sau vor scădea exponențial. Cu toate acestea, se poate demonstra că, pentru orice model de neuron, regula lui Hebb este instabilă. Prin urmare, modelele de rețele de neuroni utilizează de obicei alte teorii de învățare, cum ar fi teoria BCM, regula lui Oja sau Algoritmul Hebbian generalizat.
Vezi și
- Învățare anti-Hebbian
- Teoria BCM
- Detecția coincidențelor în neurobiologie
- Principiul lui Dale
- Algoritmul Hebbian Generalizat
- Leabra
- Lung-termen lung
- Memorie
- Metaplasticitate
- Rețele neuronale
- Regula de învățare Oja
- Stimulare tetanică
- Plasticitate dependentă de sincronizare
- Ipoteza sinaptotropică
- Shouval, Harel Fizica creierului. Bazele sinaptice ale învățării și memoriei: O abordare teoretică. The University of Texas Health Science Center at Houston. URL accesat la data de 2007-11-14.
Lecturi suplimentare
- Hebb, D.O. (1949), The organization of behavior, New York: Wiley
- Hebb, D.O. (1961). „Distinctive features of learning in the higher animal” J. F. Delafresnaye (Ed.) Brain Mechanisms and Learning, London: Oxford University Press.
- Hebb, D.O., și Penfield, W. (1940). Comportamentul uman după îndepărtarea bilaterală extinsă a lobilor frontali. Archives of Neurology and Psychiatry 44: 421-436.
- Allport, D.A. (1985). „Distributed memory, modular systems and dysphasia” Newman, S.K. și Epstein, R. (Eds.) Current Perspectives in Dysphasia, Edinburgh: Churchill Livingstone.
- Bishop, C.M. (1995). Neural Networks for Pattern Recognition, Oxford: Oxford University Press.
- Paulsen, O., Sejnowski, T. J. (2000). Modele naturale de activitate și plasticitate sinaptică pe termen lung. Opinie curentă în neurobiologie 10 (2): 172-179.
- Overview
- Hebbian Learning tutorial (Part 1: Novelty Filtering, Part 2: PCA)
|
v-d-e
Neuroetiologie |
||
|---|---|---|
| Concepte în neuroetiologie |
Feedforward – Detector de coincidențe -. Umwelt – Instinct – Detector de trăsături – Generator central de tipare (CPG) -Receptor NMDA – Inhibiție laterală – Tipare de acțiune fixă – Principiul lui Krogh-Teorie Hebbian- Localizarea sunetului |
200px
|
| Istoria neuroetologiei |
Theodore Holmes Bullock – Walter Heiligenberg -Niko Tinbergen- Konrad Lorenz- Eric Knudsen-Donald Griffin – Donald Kennedy – Karl von Frisch – Erich von Holst – Jörg-Peter Ewert |
|
| Methods in Neuroethology |
Whole Cell Patch Clamp |
|
| Model Systems in Neuroethology |
Animal Echolocation – Waggle Dance- Pești electrici – Viziunea la broaște – Neuroetologia audiției la broaște – Detecția în infraroșu la șerpi |
|
Această pagină folosește conținut licențiat Creative Commons de la Wikipedia (vezi autori).
Lasă un răspuns