Structura și funcția celulelor bacteriene
On octombrie 10, 2021 by adminStructura și funcția celulelor bacteriene (pagina 5)
(Acest capitol are 10 pagini)
© Kenneth Todar, PhD
Partea celulară
Partea celulară a bacteriilor merită o atenție specială din mai multe motive:
1. Ele sunt o structură esențială pentru viabilitate, așa cum a fost descrisă mai sus.
2. Ele sunt compuse dincomponente unice care nu se găsesc nicăieri altundeva în natură.
3. Ele sunt unul dintre cele mai importante situsuri de atac al antibioticelor.
4. Ele oferă liganzi pentru aderență și situsuri de recepție pentru medicamente sau viruși.
5. Provoacă simptome de boală la animale.
6. Asigură distincția imunologică și variația imunologică între tulpinile de bacterii.
Majoritatea procariotelor au un perete celular rigid.Peretele celular este o structură esențială care protejează protoplastul celular de deteriorarea mecanică și de ruperea sau liza osmotică.Procariotele trăiesc de obicei în medii relativ diluate, astfel încâtacumularea de substanțe solubile în interiorul citoplasmei celulei procariote depășește cu mult concentrația totală absolută din mediul exterior. Astfel, presiunea osmotică din interiorul membranei plasmatice poate fi echivalentă cu 10-25atm. Deoarece membrana este o structură delicată, plastică, ea trebuie să fie reținută de un perete exterior realizat dintr-un material poros, rigid, care are o rezistență mare la tracțiune. Un astfel de material este mureina, componenta omniprezentă a pereților celulari bacterieni.
Mureina este un tip unic de peptidoglican, un polimer de dizaharide (glican) reticulat de lanțuri scurte de aminoacizi (peptide). Există mai multe tipuri de peptidoglican. Toți peptidoglicanii bacterieni conțin acid N-acetilmuramic, care este componenta definitivă a mureinei. Pereții celulari ai Archaea pot fi compuși din proteine, polizaharide sau molecule asemănătoare peptidoglicanului, dar nu conțin niciodată mureină. Această caracteristică distinge Bacteriile de Archaea.
În bacteriile Gram-pozitive (cele care rețin colorantul violet cristal purpuriu atunci când sunt supuse procedurii de colorare Gram), peretele celular este format din mai multe straturi de peptidoglican. În mod perpendicular pe foliile de peptidoglican se află un grup de molecule numite acizi teicoici, care sunt unice pentru peretele celular Gram-pozitiv (figura 14). 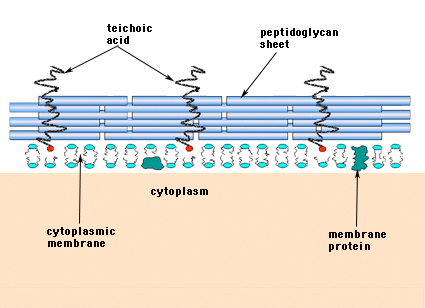
Figura 14. Structura peretelui celular al bacteriilor Gram-pozitive. Peretele este relativ gros și constă din mai multe straturi de peptidoglican întrepătrunse cu acizi teicoici care se desfășoară perpendicular pe foile de peptidoglican.
La bacteriile Gram-negative (care nu rețin cristalul violet), peretele celular estecompus dintr-un singur strat de ptidoglicani înconjurat de o structură membranoasă numită membrană externă.Membrana externă a bacteriilor Gram-negative conține invariabil uncomponent unic, lipopolizaharida (LPS sau endotoxina),care este toxic pentru animale. La bacteriile Gram-negative, membrana externă este considerată de obicei ca făcând parte din peretele celular (figura15).

Figura15. Structura peretelui celular Gram-negativ. Peretele este relativ subțire și conține mult mai puțin peptidoglican decât peretele Gram-pozitiv. de asemenea, acizii teicoici sunt absenți. Cu toate acestea, peretele celular Gram negativ este alcătuit dintr-o membrană externă care se află în afara stratului de peptidoglican. Membrana exterioară este atașată de stratul de peptidoglican prin intermediul unui grup unic de molecule de lipoproteine.
La bacteriile Gram pozitive, peretele celular este gros (15-80nanometri), format din mai multe straturi de peptidoglican. La bacteriile Gram-negative, peretele celular este relativ subțire (10 nanometri) și este alcătuit dintr-un singur strat de ptidoglican înconjurat de o membrană externă.
Structura și dispunerea peptidoglicanului la E.coli este reprezentativă pentru toate Enterobacteriaceae, precum și pentru multe alte bacterii Gram-negative. Coloana vertebrală a glicanului este alcătuită dinmolecule alternative de N-acetilglucozamină (G) și acid N-acetilmuramic (M)conectate printr-o legătură beta 1,4-glicozidică. Carbonul 3 al acidului N-acetilmuramic (M) este substituit cu o grupare lactil eter derivată din piruvat. Eterul lactilic leagă coloana vertebrală glicanică de un lanț lateral peptidic care conține L-alanină (L-ala), D-glutamat (D-glu), acid diaminopimelic (DAP) și D-alanină (D-ala). MurNAc este unic pentru pereții celulari bacterieni, la fel ca și D-glu, DAP și D-ala. Subunitatea de acid muramic din E. coli este prezentată înFigura16 de mai jos.

Figura 16. Structura subunității de acid muramic a peptidoglicanului din Escherichia coli.Acesta este tipul de mureină care se găsește în majoritatea bacteriilor Gram-negative. Coloana vertebrală a glicanului este un polimer repetat de două aminozaharuri, N-acetilglucozamina (G) și acidul N-acetilmuramic (M). De acidul N-acetilmuramic este atașată o tetrapeptidă formată din L-ala-D-glu-DAP-D-ala. b. Structura prescurtată a subunității de acid muramic. c. Lanțurile laterale tetrapeptidice apropiate pot fi legate între ele printr-o legătură interpeptidică între DAP pe un lanț și D-ala pe celălalt. d. Forma polimerică a moleculei.
Firmele de mureină sunt asamblate înperiplasmă din aproximativ 10 subunități de acid muramic. Apoi, firele sunt conectate pentru a forma o moleculă glicanică continuă care cuprinde celula.Ori de câte ori proximitatea lor o permite, lanțurile tetrapeptidice care se proiectează din coloana vertebrală glicanică pot fi reticulate printr-o legătură interpeptidicăîntre o grupare amino liberă de pe DAP și o grupare carboxi liberă de pe o D-ala din apropiere.Asamblarea peptidoglicanului în exteriorul membranei plasmatice este mediată de un grup de enzime periplasmatice, care sunt transglicozilazele, transpeptidazele și carboxipeptidazele. Mecanismul de acțiune al penicilinei și al antibioticelor beta-lactame înrudite constă în blocarea enzimelor transpeptidază și carboxipeptidază în timpul asamblării peretelui celular de mureină. Prin urmare, se spune că antibioticelebetalactamice „blochează sinteza peretelui celular” înbacterii.
Spatema glicanică a moleculei peptidoglicanului poate fi scindată de o enzimă numită lizozimă care este prezentă în serulanimal, în țesuturi și secreții și în lizozomul fagocitar. Funcția lizozimei este de a liza celulele bacteriene ca o apărare constitutivă împotriva agenților patogeni bacterieni. Unele bacterii Gram-pozitive sunt foarte sensibile la lizozimă, iar enzima este destul de activă la concentrații mici.Secrețiile lacrimale (lacrimi) pot fi diluate 1:40.000 și își păstrează capacitatea de a liza anumite celule bacteriene. Bacteriile Gram-negative sunt mai puțin vulnerabile la atacul lizozimei, deoarece peptidoglicanul lor este protejat de membrana exterioară. Locul exact de scindare a lizozimei este legătura beta 1,4 dintre acidul N-acetilmuramic (M) și N-acetilglucozamina (G), astfel încât subunitatea de aciduramic prezentată în figura 16(a) este rezultatul acțiunii lizozimei asupra peptidoglicanului bacterian.
La bacteriile Gram-pozitive există numeroase aranjamente peptidice diferite între peptidoglicani. Cel mai bine studiat este cel alureino de Staphylococcus aureus prezentat în figura 17 de mai jos. În locul DAP (la E. coli) se află diaminoacidul, L-lizina (L-lys), iar în locul legăturii interpeptidice (la Gram-negative) se află o punte interpeptidică de aminoacizi care leagă o grupare amino liberă de pe lizină de o grupare carboxi liberă de pe D-ala a unui lanț lateral tetrapeptidic apropiat. Această organizare permite, aparent, o legătură încrucișată mai frecventă între catenele laterale tetrapeptidice apropiate. La S. aureus, puntea interpeptidică este o peptideconstituită din 5 molecule de glicină (numită punte pentaglicină). Asamblarea punții interpeptidice în mureina Gram-pozitivă este inhibată de antibioticele beta lactamice în același mod ca și legătura interpeptidică în mureina Gram-negativă. Bacteriile Gram-pozitive sunt mai sensibile la penicilină decât cele Gram-negative deoarece peptidoglicanul nu este protejat de o membrană exterioară și este o moleculă mai abundentă. La bacteriile Gram-pozitive, peptidoglicanii pot varia în ceea ce privește aminoacidul din locul DAP sau L-lys din poziția 3 a tetrapeptidei și în ceea ce privește compoziția exactă a punții interpeptidice. Cel puțin opt tipuri diferite de peptidoglicani există în bacteriile Gram-pozitive.
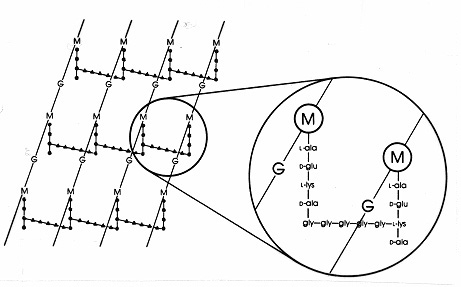
Figura 17. Diagramă schematică a foliei de peptidoglican a Staphylococcus aureus. G = N-acetil-glucozamină;M = acid N-acetil-muramic; L-ala = L-alanină; D-ala = D-alanină; D-glu= acid D-glutamic; L-lys = L-lizină. Acesta este un tip de mureină care se găsește în bacteriile Gram-pozitive. Comparativ cu peptidoglicanul din E. coli (figura 7), în locul DAP (acidul diaminopimelic) din tetrapeptidă există L-lys.Grupa amino liberă a L-lys este înlocuită cu o glicinepentapeptidă (gly-gly-gly-gly-gly-gly-gly-gly-) care devine apoi o punte interpeptidică formând o legătură cu o grupare carboxi din D-ala într-o catenă laterală tetrapeptidică adiacentă. Peptidoglicanii Gram-pozitivi diferă de la o specie la alta, în principal în ceea ce privește aminoacizii din a treia poziție a lanțului tetrapeptidic și în compoziția de aminoacizi a punții interpeptidice.
Bacteriile Gram-negative pot conține un singur strat monomolecular de mureină în pereții lor celulari, în timp ce bacteriile Gram-pozitive sunt considerate ca având mai multe straturi sau „învelișuri” de peptidoglican. La bacteriile Gram-pozitive, straturile de peptidoglican sunt strâns asociate cu un grup de molecule numite acizi teicoici. Acizii teicoici sunt polimeri liniari de poliglicerol sau polibitol substituiți cu fosfați și câțiva aminoacizi și zaharuri. Polimerii acizilor teicoici sunt ocazional ancorați la membrana plasmatică (numiți acid lipoteicoichoic, LTA), aparent orientați spre exterior, în unghiuri drepte față de straturile de peptidoglican. Funcțiile acidului teicoichoic nu sunt cunoscute. Acestea sunt esențiale pentru viabilitatea bacteriilor Gram-pozitive în mediul natural. O idee este aceea că acestea oferă un canal de sarcini negative orientate în mod regulat pentru trecerea substanțelor încărcate pozitiv prin rețeaua complicată de peptidoglican. O altă teorie este aceea că acizii teicoici sunt implicați într-un fel în reglarea și asamblarea subunităților de acizi muramici în exteriorul membranei plasmatice. Există cazuri, în special la streptococi, în care acizii teicoici au fost implicați în aderența bacteriilor la suprafețele țesuturilor.
Capitolul continuă
Prima pagină
.
Lasă un răspuns