Frontiers in Psychology
On septembrie 28, 2021 by adminCorpul calos (CC) este comisura primară care conectează cele două emisfere cerebrale ale creierului. Prin urmare, deși termenul de comisurotomie nu este incorect pentru a descrie așa-numitul „split-brain” și este strict corect pentru operațiile chirurgicale originale la om în care toate comisurile au fost secționate (Gazzaniga et al., 1967), intervențiile chirurgicale ulterioare au secționat doar CC, aducând astfel în uz termenul de „callosotomie”, care rămâne ca termen mai adecvat în aceste cazuri (de exemplu, Franz et al., 1996, 2000).
Dintr-o perspectivă evolutivă, CC este o dezvoltare recentă observată doar la mamiferele placentare (Katz et al., 1983; Mihrshahi, 2006). Se crede că funcționalitatea asigurată de CC a fost precedată, cel puțin parțial, de comisura anterioară (CA), așa cum se observă la mamiferele neplacentare, cum ar fi marsupialele (Heath și Jones, 1971; Ashwell et al., 1996). Deși unele căi reziduale ale CA sunt încă prezente în creierul uman, conectivitatea și structura sa rămân insuficient studiate în literatura actuală, iar constatările rare sunt mai degrabă inconsistente (Lasco et al., 2002; Patel et al., 2010; Choi et al., 2011). De exemplu, constatările bazate pe imagistica prin rezonanță magnetică (IRM) sunt amestecate în ceea ce privește faptul dacă CA are dimensiuni similare la femei și bărbați (Lasco et al., 2002; Patel et al., 2010). Într-un studiu relativ recent care utilizează tehnica in-vivo a imagisticii tensorului de difuzie (DTI), care valorifică difuzia moleculelor de apă în axoni, Patel et al. (2010) au studiat opt subiecți și au izolat cu succes fibrele CA la cinci subiecți. Printre constatări s-au numărat, la un subiect, o proporție mare de fibre din membrul posterior al AC care călătoreau bilateral în regiunile parietale, unde cele mai multe dintre ele se terminau, cu un fascicul mai mic care se termina în girusul precentral și încă un fascicul care intra în lobul temporal. Astfel de diferențe individuale au fost găsite la toți participanții, totuși majoritatea subiecților au prezentat proiecții bilaterale către cortexul orbitofrontal, parietal și temporal, în care CC nu se proiectează în mod clar. Împreună, constatările implică faptul că CA are atât proiecții primare, cât și vestigiale către zonele neocorticale în care CC nu se proiectează, așa cum au sugerat și alții (Di Virgilio et al., 1999; Patel et al., 2010).
În opinia noastră, AAA se bazează foarte mult pe rețelele neocorticale și pe conectivitatea cu structuri subcorticale, cum ar fi ganglionii bazali și talamusul (printre altele care nu sunt discutate în mod specific în propunerea noastră) (Franz, 2012) (Franz, 2012). Astfel, orice evoluție evolutivă care implică AC ar putea avea, de asemenea, un anumit rol în modelul AAA. Cu toate acestea, am găsit puțină literatură care să abordeze în mod specific această posibilă legătură.
Rolul AC a fost studiat în sarcini care sunt susceptibile de a necesita forme de atenție, cum ar fi cele care implică atenția vizuală. De exemplu, este bine cunoscut faptul că populațiile acallosale au fost studiate în contextul sarcinilor care implică integrarea vizuală (Gazzaniga et al., 1962, 1965; Sperry, 1968; Levy et al., 1972). Mai recent, Corballis (1995) a căutat să abordeze integrarea vizuală și atenția într-un efort de a explica discrepanțele anterioare în cercetarea acallosală. Corballis a propus un sistem de atenție vizuală dihotomic; o parte (implicând forma) este automată și localizată în fiecare emisferă, iar cealaltă (implicând mișcarea și localizarea) este voluntară și subcorticală. Modelul AAA se poate baza pe această observație și poate aborda discrepanțele observate în literatura de specialitate. Pe baza modelului nostru propus, pe mai multe niveluri de atenție există și alte căi decât CC care ar putea media transferul de informații între emisfere. O posibilitate implică circuitele ganglionilor bazali, pe care noi le propunem ca fiind potențial critice în medierea partajării atenției între emisfere (Franz, 2012). Prin secționarea CC, transferul direct al atenției prin intermediul căilor corticale este în mare măsură abolit (cu excepția cazului în care, desigur, CA joacă un rol de rezervă). Cu toate acestea, integrarea bazată pe atenție a informațiilor vizuale ar putea fi în continuare mediată prin intermediul căilor subcorticale și/sau al CA. În acest mod, sistemul de atenție în două părți observat de Corballis (1995) ar putea fi prezent datorită medierii bazate pe atenție a integrării vizuale prin intermediul AC (și/sau ganglionii bazali și circuitele implicate: Franz, 2012).
Studiile care implică viziunea și atenția nu trebuie să fie considerate în întregime separat de sistemul care reglează acțiunea motorie. Pe baza propriilor noastre constatări (Franz, 2004; Franz și Packman, 2004), diverse forme de atenție (vizuală și internă-nonvizuală) sunt strâns legate de acțiunea motorie și este logic că o posibilă cheie pentru înțelegerea unui potențial rol de atenție al AC se află în legătura dintre acțiune, viziune și atenție. Într-un studiu anterior, Franz (2004) a pus participanții normali să deseneze cercuri bimanuale sub diferite manipulări ale feedback-ului vizual, sau ale atenției interne la fiecare mână. Dimensiunea cercurilor desenate de o mână care primea o formă de atenție internă a devenit mai mare într-un mod similar cu efectele care au avut loc în prezența feedback-ului vizual al acelei mâini (în raport cu lipsa de atenție/backback vizual). Modelul nostru AAA implică atenția ca fiind o componentă cheie în reglarea ieșirii motorii (precum și în selectarea acțiunii motorii planificate) și, în măsura în care dovezile prezente, nu putem exclude faptul că AC joacă un rol în această funcție. Cu toate acestea, testarea directă a rolului CA în orice sarcină cognitivă nu este trivială.
Testarea dacă CA participă la funcția cognitivă este aproape imposibilă în cercetarea neuropsihologică, având în vedere că participanții cu CA localizat nu sunt ușor de găsit (și nu cunoaștem niciunul). Prin urmare, ar putea fi mult mai intuitiv să se evalueze diferențele neurologice în CA normal și să se coreleze datele neurologice cu datele comportamentale. Metoda de tractografie prezentată (elucidarea AC) poate fi aplicată relativ ușor pe eșantioane mari de populație normală și, în mod ideal, poate fi combinată cu performanțele (bazate pe sarcini efectuate în timpul unor sesiuni de testare separate de colectarea DTI) ale acelorași indivizi folosind sarcini cognitive concepute în acest scop. În prezenta lucrare, demonstrăm că DTI poate fi efectuată și poate produce rezultate utilizabile pe un eșantion mic de persoane din populația normală din punct de vedere neurologic. Propunerea generală ar fi să se efectueze metode similare pe un eșantion mai mare și (așa cum s-a sugerat mai sus), să se coreleze rezultatele (ale unei măsuri proxy a densității CA, de exemplu) cu variabilele dintr-o sarcină cognitivă.
În Figura 1 prezentăm rezultatele urmăririi DTI pe date obținute de la 10 participante de sex feminin obținute dintr-un set de date DTI publice de la Universitatea Johns Hopkins (a se vedea Recunoștințe pentru sursa completă). Vârsta medie a fost de 22 de ani (SD = 2,5). Imaginile au fost procesate folosind FSL Diffusion Toolbox (Smith et al., 2004; Woolrich et al., 2009; Jenkinson et al., 2012), iar procedurile au folosit cele utilizate în studiile anterioare pe alte trasee de fibre (FDT; Behrens et al., 2003a,b, 2007; Johansen-Berg et al., 2004). Pe scurt, imaginile au fost mai întâi concatenate în volume 4d pentru fiecare participant și craniile au fost decapate folosind instrumentul Brain Extraction Tool (BET; Smith, 2002; Jenkinson și colab., 2005) al FSL. Imaginile extrase au fost corectate pentru mișcarea minoră a capului și curenții turbionari generați de schimbările în direcția câmpului. Tensorii de difuzie au fost ajustați pentru a genera imagini care afișează direcțiile primare de difuzie și nivelurile de FA la fiecare voxel pentru a fi utilizate pentru hărțile de culori în timpul mascării. Estimarea bayesiană a parametrilor de difuzie obținuți folosind tehnici de eșantionare cu fibre de încrucișare (BEDPOSTX; „X” implicând fibre de încrucișare) a fost realizată pentru a genera o distribuție a mărimii și a direcției de difuzie care să țină cont de două fibre de încrucișare pentru a efectua ulterior tractografia pe voxeli de difuzie.
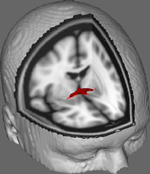
Figura 1. Rezultatele medii de urmărire la 10 participanți de control suprapuse peste o imagine IRM standard ponderată T1 cu voxeli izotropi de 2 mm pentru referință anatomică.
Rezultatele medii de urmărire la toți cei 10 participanți de control au fost apoi suprapuse peste o imagine IRM standard ponderată T1 cu voxeli izotropi de 2 mm pentru referință anatomică. Raportul FA mediu al grupului a fost de 0,3146 (SD = 0,0472). Aceste rezultate ale tractografiei arată, în mod important, că structura AC poate fi elucidată și, prin urmare, are potențial pentru studii ulterioare care să coreleze scorurile de performanță pe sarcini comportamentale cu valorile FA folosind o dimensiune mai mare a eșantionului.
În rezumat, în prezent, există o literatură rară, iar din literatura disponibilă, niciuna nu leagă în mod specific AC de procesele legate de modelul AAA, care este punctul central al articolului nostru țintă anterior (Franz, 2012). Folosind o procedură similară cu cea propusă aici, este posibil să se testeze eșantioane mai mari de populație normală și să se coreleze valorile FA ale rezultatelor DTI (de exemplu) cu o variabilă dependentă specifică obținută dintr-o sarcină de atenție la acțiune dezvoltată în mod intenționat (ale cărei proceduri sunt o dezvoltare în curs de desfășurare în laboratorul nostru).
Declarație privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Recunoștințe
Recunoaștem cu recunoștință Human Brain Project și National Research Resource Center (SUA) și Universitatea John Hopkins pentru că au permis utilizarea publică a datelor lor DTI. Suntem, de asemenea, recunoscători pentru sprijinul financiar generos din partea Fondului Marsden al Societății Regale din Noua Zeelandă (pentru Elizabeth A. Franz).
Corballis, M. (1995). Integrarea vizuală în creierul divizat. Neuropsychologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). Alocarea atenției pentru învățarea acțiunilor orientate spre scopuri: un cadru de neuroștiință cognitivă care se concentrează pe ganglionii bazali. Front. Psychol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | Refef Full Text
Heath, C., and Jones, E. (1971). Căile interhemisferice în absența unui corp calos. Un studiu experimental al conexiunilor comisurale la falanga marsupială. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
.
Lasă un răspuns