Tydzień Rekina: What Did Megalodon Eat?
On 13 grudnia, 2021 by adminGuest Author: Jack Cooper, MSc
UoB Graduate / PhD Student, University of Swansea
Jak widzieliśmy do tej pory, następowały zmiany i aktualizacje naukowego konsensusu co do taksonomicznego przydziału Megalodona i maksymalnego rozmiaru ciała. Jednakże, jego ulubiona przekąska jest czymś, co jest powszechnie uzgodnione w społeczności naukowej. Co więcej, jest to coś, co zazwyczaj jest dość dokładnie przedstawiane w literaturze pięknej. Pozwólcie, że przedstawię rekina, który jadł wieloryby – prawdziwe wieloryby!
Wiele dzisiejszych dużych rekinów drapieżnych jest znanych z jedzenia ssaków morskich, zwłaszcza nasz stary przyjaciel żarłacz biały. Niektóre z najbardziej kultowych zdjęć tego rekina przedstawiają go wynurzającego się z wody z ostatnią ofiarą w szczękach (ryc. 1). Młodociane żarłacze białe nie zjadają jednak fok od samego początku. Zamiast tego, zaczynają od jedzenia ryb i mniejszych rekinów. Jeśli spojrzymy ponownie na formację Gatun w Panamie, znajdziemy mnóstwo skamieniałych rekinów, płaszczek i otolitów (struktur z uszu wewnętrznych ryb), a także mnóstwo bezkręgowców, takich jak mięczaki .
Częścią, której jednak nie znajdujemy, są skamieniałe ssaki morskie. To wydaje się wskazywać, że Megalodon młodych były również jedzenie ryb i mniejszych rekinów, a nie od razu iść po ssaków morskich. Jest to coś, co nazywa się ontogenetyczną zmianą diety, co oznacza, że rekin zmienia preferowane źródło pożywienia w miarę wzrostu. Jedząc większe pokarmy, i prawdopodobnie dużo, jak osiągnął swój dorosły rozmiar, Megalodon byłby w stanie utrzymać metaboliczne wymagania bycia tak dużym. Mógł to zrobić jedząc nie tylko wieloryby, ale po prostu wszystko, na co się natknął.

Given ile jedzenia Megalodon musiałby zjeść, aby uzasadnić swój ogromny rozmiar, prawie na pewno był oportunistycznym karmicielem. Zapis kopalny jest tu naszym najlepszym przewodnikiem. Wynika to z obecności skamieniałości śladowych, które są geologicznymi zapisami aktywności biologicznej. Mogą to być odciski stóp dinozaurów po zadrapania na kościach wskazujące, że coś ugryzło zwierzę, do którego należała kość. A Megalodon pozostawił kilka takich skamieniałości śladowych za sobą, dostarczając kluczowych dowodów swoich zabójstw.
Znaleziono kilka skamieniałości należących do różnych waleni z godnymi uwagi śladami ugryzień na nich. Jeden dobrze cytowany przykład pochodzi z formacji Yorktown w Maryland, gdzie kilka kości wielorybów miało ślady ugryzień, które zostały wyraźnie wykonane przez duże ząbkowane zęby. W rzeczywistości, zęby Megalodon są czasami znaleźć bezpośrednio związane z tymi kośćmi .
Rekiny tracą i zastępują zęby tysiące razy w ciągu swojego życia, a zatem rekiny często tracą zęby podczas dokonywania gwałtownego zabójstwa. Jest to kluczowy powód, dla którego zęby rekinów są tak powszechnymi skamieniałościami. Obecność zębów rekina tuż przy porysowanej kości wieloryba wskazuje na to, że żerowanie miało miejsce. Jednak to, co paleontolodzy powinni zawsze brać pod uwagę, gdy znajdują takie skamieniałości, to pytanie: czy było to rzeczywiście drapieżnictwo? A może było to padlinożerstwo?
Padlinożerstwo jest powszechnym zachowaniem żywieniowym u współczesnych rekinów drapieżnych, takich jak żarłacze białopłetwe (ryc. 2) i rekiny tygrysie. Kiedy rekiny te natrafiają na unoszące się na powierzchni martwe wieloryby, jest to bufet, w którym wszystko można zjeść. Co ważne, padlinożerstwo nie wymaga wydatkowania energii, która może zostać zmarnowana podczas nieudanej próby drapieżnictwa. Biorąc pod uwagę, że Megalodon musiał być w stanie utrzymać swoje ogromne rozmiary, nie miałoby sensu rezygnować z darmowego posiłku, gdyby natknął się na martwego wieloryba. Tak więc, na podstawie tego, jak duże współczesne rekiny się odżywiają, większość z nas byłaby skłonna założyć się, że Megalodon prawdopodobnie padlinożerny, gdy miał taką możliwość.

To powiedział, jest jeden szczególnie niezwykły skamielina, która implikuje Megalodon w próbie drapieżnictwa. I, co istotne, wiemy to przede wszystkim dlatego, że próba ta zakończyła się niepowodzeniem. W artykule opublikowanym w 2010 roku opisano skamielinę części żebra z Pliocenu należącą do walenia z formacji Yorktown w Karolinie Północnej (tak, dwie formacje Yorktown). Skamieniałość ta również wykazywała uszkodzenia przypominające ślady ugryzień, ale co istotne, na ślady te nakładała się tkana kostna powłoka. Autorzy zinterpretowali to jako oznakę infekcji, która pojawiła się, gdy zwierzę dochodziło do siebie po urazie kości spowodowanym przez te ślady ugryzień. Ślady te dobrze pasują do ząbkowanego zgryzu, co sugeruje, że napastnik miał ząbkowane zęby – tak jak Megalodon. Badanie sugerowało więc, że Megalodon, lub inny duży rekin, zaatakował zwierzę, z którego pochodziła skamielina, ale ofiara przeżyła próbę tylko po to, by sześć tygodni później umrzeć z powodu infekcji wywołanej obrażeniami.
Nawet ostatnio, pierwsze w historii kopalne ślady ugryzień znalezione na półkuli południowej, przypisywane Megalodonowi, znaleziono w peruwiańskiej formacji Pisco. Być może tak miało być, gdyż jest to to samo miejsce, w którym Gordon Hubbell znalazł swoją wyjątkową skamieniałość Carcharodona (omówioną w Q1). Znaleziony tu materiał zawierał szczątki czaszki i żeber, co sugeruje, że rekin ugryzł swój cel z boku. Co najważniejsze, szczątki te zostały po raz pierwszy zidentyfikowane jako osobny gatunek – tym gatunkiem jest Piscobalaena nana, mały wieloryb łuskopodobny. Podobnie jak w przypadku innych skamieniałości śladowych, ślady ugryzień dobrze pasowały do dużych ząbkowanych zębów – co przemawia za oskarżeniem Megalodona o atak (ryc. 3). Okazało się zatem, że Megalodon był również zdolny do atakowania mniejszych wielorybów (czy to w wyniku drapieżnictwa, czy padlinożerstwa), co jest zgodne z sugestiami, że jego celem były prawdopodobnie również delfiny, żółwie morskie i inne mniejsze ofiary

Choć strategie drapieżnicze są trudne do zinterpretowania na podstawie skamieniałości, współczesne rekiny mogą dać nam wyobrażenie o tym, jak polował Megalodon. Jego siła ugryzienia byłaby wyjątkowo potężna do uszkadzania kości wielorybów. Komputerowy model czaszki żarłacza białego wykazał, że jego siła ugryzienia przekracza około 1,8 tony, co daje mu jedną z najsilniejszych sił ugryzienia wśród wszystkich żyjących zwierząt. Rozszerzając, to badanie sugeruje, że siła ugryzienia Megalodona była jeszcze większa, szacując na niezwykłe 108,514-182,201 N.
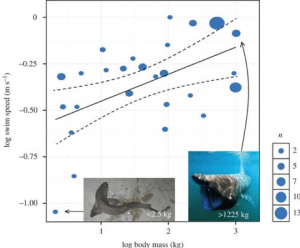
Naukowcy podjęli również próby obliczenia prędkości pływania Megalodona, co może pomóc dać nam wskazówkę, jak polował. Rekiny mogą mieć prędkość zrywu, która jest szybkim wstrząsem szybkiego pływania, zazwyczaj używanym do chwytania zdobyczy. Jednak ich zwykła prędkość podczas swobodnego pływania jest tym, co nazywamy stałą prędkością pływania. Różni się to w zależności od gatunku, ale jedno z badań pod kierownictwem Davida Jacoby’ego połączyło dane dotyczące 26 gatunków z 64 wcześniejszych badań, aby opracować model, który przewidywałby zależność skalowania pomiędzy stałą prędkością pływania a metabolizmem i masą ciała rekina. Ich model uwzględniający metabolizm ujawnił wykładnik skalowania równy 0,173, podsumowany następującym równaniem:
Szybkość pływania (ms ¹) ∝ Masa (kg)^0,173
Kiedy jednak skorygowano dane o filogenezę, okazało się, że minimalna prędkość pływania skaluje się z masą ciała z nieco niższym wykładnikiem 0,15. Powyższe równanie zakłada dodatkowo stałą równą 1, aby dopasować proporcje, ale dane ujawniły, że stała ta wynosiła w rzeczywistości 0,266. Dlatego skonstruowali równanie mocy:
Szybkość pływania (ms ¯¹) = 0,266 Masa (kg) ^0,15
Z tego są w stanie obliczyć trwałą prędkość pływania Megalodona w zależności od jego masy. Na przykład Megalodon o masie ~48 000 kg, obliczony w pracy Gottfrieda jako masa rekina o długości 15,9 m, ujawniłby rekina pływającego nieprzerwanie z prędkością 1,34 m/s – około 4,8 km/godzinę. Zastosowanie tego do Megalodona o masie 52 000 kg ujawniło stałą prędkość pływania 1,36 m/s lub 4,9 km/godz. Gdybyśmy jednak popadli w skrajność i symulowali rekina o masie 100 000 kg, model ten wykazałby prędkość pływania 1,50 m/s lub 5,4 km/godz. Model ten okazał się bardzo skuteczny nie tylko w obliczaniu prędkości pływania Megalodona, ale także w dostarczaniu modelu dla wielu gatunków rekinów (ryc. 4) .
Jednakże to właśnie prędkość zrywu rekiny wykorzystują do szybkiego pochwycenia ofiary. Żarłacze białe wykorzystują to w swoim „breaching behaviour”, w którym napędzają się z wody, pędząc ofiarę od dołu, aby złapać ją z zaskoczenia (Rys. 1) . Mogą to robić, ponieważ są dobrze zakamuflowane dzięki przeciwzaciemnieniu. Tak więc, ponieważ Megalodon był prawdopodobnie również zakamuflowany i miał podobną ekologię i budowę ciała do żarłaczy białych, nasuwa się pytanie: czy Megalodon był również zdolny do rozerwania się?
Możliwe. Prędkość uderzeniowa żarłacza białego jest napędzana przez fizjologiczną adaptację zwaną mezotermią. Jest to coś, co Megalodon niedawno zasugerowano, że również posiadał (zostanie to szczegółowo omówione w Q5). Mezotermiczne ryby ogólnie mają szybsze trwałe i burst prędkości pływania niż ektotermiczne ryby , a to zostało wykorzystane do sugerowania, że Megalodon może mieć prędkość burst do 37.15 km / h .
Ten niewiarygodnie szybki prędkość dla takiego dużego rekina może być wystarczające, aby być w stanie naruszyć. Poza tym, rekin olbrzym (Cetorhinus maximus), dziś drugi co do wielkości rekin, również został zaobserwowany. Być może gigantyczny Megalodon rzeczywiście był w stanie wyskoczyć z wody, gdy chwytał swój najnowszy posiłek. Po prostu nie wiemy jeszcze, ale moja osobista i (nieco) profesjonalna opinia jest taka, że mogło to być możliwe.
Z tak szerokim menu do wyboru, Megalodon był drapieżnikiem wierzchołkowym, bez znanych drapieżników własnych. Oznacza to dwie ważne rzeczy. Po pierwsze, prawdopodobnie miał silną konkurencję dla swoich ofiar, coś, co mogło ostatecznie przyczynić się do jego wyginięcia. Po drugie, jako drapieżnik wierzchołkowy, Megalodon byłby niezwykle ważny dla swojego ekosystemu, ponieważ utrzymałby populacje ofiar w ryzach, a tym samym zachował zdrowy ekosystem.
To dlatego dzisiejsze rekiny są tak ważne dla środowisk morskich. Utrata rekina drapieżnego apex produkuje nieodwracalne efekty kaskadowe, które zmieniają społeczność, którą pozostawia za sobą. Jako takie, nasze ekosystemy morskie są takie, jakie są częściowo dlatego, że Megalodon nie jest już ich częścią. A ogromne wieloryby współczesnych oceanów mogą w rezultacie spać spokojnie.
- Estrada JA, Rice AN, Natanson LJ & Skomal GB 2006. Zastosowanie analizy izotopowej kręgów w rekonstrukcji ontogenetycznej ekologii żywienia u żarłaczy białych. Ecology 87, 829-834.
- Pimiento C, Ehret DJ, MacFadden BJ & Hubbell G 2010. Ancient nursery area for the extinct giant shark Megalodon from the Miocene of Panama. PLoS One 5, e10552.
- Pimiento C, González-Barba G, Ehret DJ, Hendy AJ, MacFadden BJ & Jaramillo C 2013. Sharks and rays (Chondrichthyes, Elasmobranchii) from the late Miocene Gatun formation of Panama. J. Paleontol. 87, 755-774.
- Purdy RW 1996. Paleoecology of fossil white sharks. In: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 67-78, Academic Press, San Diego.
- Aguilera OA, García L & Cozzuol MA 2008. Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation. Paläontol. Z. 82, 204-208.
- Fallows C, Gallagher A. & Hammerschlag N 2013. White sharks (Carcharodon carcharias) scavenging on whales and its potential role in further shaping the ecology of an apex predator. PLoS One, 8, e60797.
- Dudley SFJ, Anderson-Reade MD, Thompson GS & McMullen PB 2000. Concurrent scavenging of a whale carcass by great white sharks, Carcharodon carcharias, and tiger sharks, Galeocerdo cuvier. Fish. Bull. 98, 646-649.
- Kallal RJ, Godfrey SJ & Ortner DJ 2010. Bone reactions on a Pliocene cetacean rib indicate short-term survival of predation event. Int. J. Osteoarchaeol. 22, 253-260.
- Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M & Bianucci G 2017. Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru. Palaeogeogr. Palaeoclimatol. Palaeoecol. 469, 84-91.
- Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, Ferrara TL, Cunningham E, Dean MN & Summers AP 2008. Trójwymiarowa analiza komputerowa mechaniki szczęki żarłacza białego: jak mocno może ugryźć wielki biały? J. Zool. 276, 336-342.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2015. Is the scaling of swim speed in sharks driven by metabolism? Biol. Lett. 11, 20150781.
- Gottfried MD, Compagno LJV & Bowman SC 1996. Size and skeletal anatomy of the giant „megatooth” shark Carcharodon megalodon. In: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 55-66, Academic Press, San Diego.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2016. Correction to 'Is the scaling of swim speed in sharks driven by metabolism?’ Biol. Lett. 12, 20160775.
- Watanabe YY, Payne NL, Semmens JM, Fox A & Huveneers C 2019. Swimming strategies and energetics of endothermic white sharks during foraging (Strategie pływania i energetyka endotermicznych żarłaczy białych podczas żerowania). J. Exp. Biol. 222, jeb185603.
- Cooper JA, Pimiento C, Ferrón HG & Benton MJ (in press). Body dimensions of the extinct giant shark Otodus megalodon: A 2D reconstruction. Sci. Rep.
- Watanabe YY, Goldman KJ, Caselle JE, Chapman DD & Papastamatiou YP 2015. Comparative analyses of animal-tracking data reveal ecological significance of endothermy in fishes. Proc. Nat. Acad. Sci. 112, 6104-6109.
- Ferrón HG 2017. Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks. PLoS One, 12, e0185185.
- Johnston EM, Halsey LG, Payne NL, Kock AA, Iosilevskii G, Whelan B & Houghton JD 2018. Latent power of basking sharks revealed by exceptional breaching events. Biol. Lett. 14, 20180537.
- Myers RA, Baum JK, Shepherd TD, Powers SP & Peterson CH 2007. Cascading effects of the loss of apex predatory sharks from a coastal ocean. Science, 315, 1846-1850.
Edited by Rhys Charles
.
Dodaj komentarz