Teoria hebrajska
On 14 grudnia, 2021 by admin Ocena | Biopsychologia | Porównawcza | Poznawcza | Rozwojowa | Językowa | Różnice indywidualne | Osobowość | Filozoficzna | Społeczna |
Metody | Statystyczna | Kliniczna | Edukacyjna | Przemysłowa | Zawodowa | Psychologia świata |
Psychologia poznawcza:Uwaga -Podejmowanie decyzji -Uczenie się -Sądzenie -Pamięć -Motywacja -Spostrzeganie -Rozumowanie -Myślenie -Procesy poznawczePoznanie -OutlineIndex
Teoria Hebba opisuje podstawowy mechanizm plastyczności synaptycznej, w którym wzrost skuteczności synaptycznej wynika z powtarzającej się i uporczywej stymulacji komórki presynaptycznej przez komórkę postsynaptyczną. Wprowadzona przez Donalda Hebba w 1949 roku, nazywana jest również regułą Hebba, postulatem Hebba, teorią składania komórek i stwierdza:
Załóżmy, że uporczywość lub powtarzanie aktywności pogłosowej (lub „śladu”) ma tendencję do wywoływania trwałych zmian komórkowych, które zwiększają jej stabilność…. Kiedy akson komórki A jest wystarczająco blisko, aby pobudzić komórkę B i wielokrotnie lub wytrwale bierze udział w jej odpalaniu, jakiś proces wzrostu lub zmiana metaboliczna zachodzi w jednej lub obu komórkach, tak że efektywność A, jako jednej z komórek odpalających B, jest zwiększona.
Teoria jest często podsumowywana jako „komórki, które odpalają razem, drut razem”, chociaż jest to zbytnie uproszczenie układu nerwowego, które nie powinno być brane dosłownie, jak również nie dokładnie reprezentuje oryginalne oświadczenie Hebba na temat zmian siły połączenia komórek. Teoria ta jest powszechnie przywoływana w celu wyjaśnienia niektórych rodzajów uczenia asocjacyjnego, w których jednoczesna aktywacja komórek prowadzi do wyraźnego wzrostu siły synaptycznej. Takie uczenie jest znane jako uczenie hebrajskie.
Hebbowskie engramy i teoria łączenia komórek
Teoria hebrajska dotyczy tego, jak neurony mogą się łączyć, by stać się engramami. Teorie Hebba na temat formy i funkcji zespołów komórek można zrozumieć z następujących:
„Ogólna idea jest stara, że jakiekolwiek dwie komórki lub systemy komórek, które są wielokrotnie aktywne w tym samym czasie, będą miały tendencję do stawania się 'stowarzyszonymi’, tak że aktywność w jednej ułatwia aktywność w drugiej.” (Hebb 1949, str. 70) „Kiedy jedna komórka wielokrotnie pomaga w odpaleniu drugiej, akson pierwszej komórki rozwija gałki synaptyczne (lub powiększa je, jeśli już istnieją) w kontakcie z somą drugiej komórki.” (Hebb 1949, str. 63)
Gordon Allport przedstawia dodatkowe idee dotyczące teorii łączenia komórek i jej roli w formowaniu engramów, zgodnie z koncepcją autoasocjacji, opisaną w następujący sposób:
„Jeśli wejścia do systemu powodują, że ten sam wzorzec aktywności występuje wielokrotnie, zestaw aktywnych elementów tworzących ten wzorzec będzie coraz silniej powiązany. To znaczy, każdy element będzie miał tendencję do włączania każdego innego elementu i (z ujemnymi wagami) do wyłączania elementów, które nie stanowią części wzorca. Innymi słowy, wzorzec jako całość stanie się „auto-zasocjowany”. Wyuczony (auto-asocjacyjny) wzorzec możemy nazwać engramem.” (Hebb 1949, s. 44)
Teoria hebrajska była podstawową podstawą konwencjonalnego poglądu, że gdy analizuje się je z poziomu holistycznego, engramy są siatkami neuronowymi lub sieciami neuronowymi.
Praca w laboratorium Erica Kandela dostarczyła dowodów na zaangażowanie mechanizmów hebrajskiego uczenia się przy synapsach u ślimaka morskiego Aplysia californica.
Experiments na Hebbian mechanizmów modyfikacji synapsy w centralnym układzie nerwowym synapsy kręgowców są znacznie trudniejsze do kontroli niż są eksperymenty z stosunkowo proste synapsy obwodowego układu nerwowego studiował w morskich bezkręgowców. Duża część prac nad długotrwałymi zmianami synaptycznymi między neuronami kręgowców (takimi jak potencjacja długotrwała) polega na wykorzystaniu niefizjologicznej stymulacji eksperymentalnej komórek mózgowych. Jednakże, niektóre z fizjologicznie istotnych mechanizmów modyfikacji synaps, które były badane w mózgach kręgowców, wydają się być przykładami procesów Hebbiana. W jednym z takich badań dokonano przeglądu wyników eksperymentów, które wskazują, że długotrwałe zmiany w sile synaptycznej mogą być wywołane przez fizjologicznie istotną aktywność synaptyczną działającą zarówno poprzez mechanizmy hebrajskie, jak i niehebrajskie
Zasady
Z punktu widzenia sztucznych neuronów i sztucznych sieci neuronowych, zasadę Hebba można opisać jako metodę określania sposobu zmiany wag pomiędzy modelowymi neuronami. Waga pomiędzy dwoma neuronami wzrasta, jeśli te dwa neurony aktywują się jednocześnie, a maleje, jeśli aktywują się osobno. Węzły, które mają tendencję do bycia pozytywnymi lub negatywnymi w tym samym czasie mają silne wagi pozytywne, podczas gdy te, które mają tendencję do bycia przeciwnymi mają silne wagi negatywne.
Ta oryginalna zasada jest być może najprostszą formą selekcji wag. Oznacza to, że można ją stosunkowo łatwo zakodować w programie komputerowym i wykorzystać do aktualizacji wag w sieci, ale jednocześnie ogranicza liczbę zastosowań uczenia hebrajskiego. Obecnie termin uczenie Hebbiana odnosi się do pewnej formy matematycznej abstrakcji oryginalnej zasady zaproponowanej przez Hebba. W tym sensie uczenie hebrajskie polega na dostosowywaniu wag między węzłami uczącymi się w taki sposób, aby każda waga lepiej odzwierciedlała relacje między węzłami. W związku z tym wiele metod uczenia się można uznać za w pewnym stopniu hebrajskie w naturze.
Poniżej znajduje się wzór opisu uczenia hebrajskiego: (zauważ, że wiele innych opisów jest możliwych)
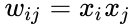
gdzie 
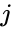
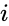
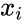
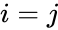
Innym wzorem opisu jest:
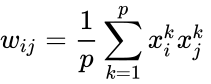
gdzie 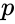

Odmianą uczenia Hebbiana, która uwzględnia takie zjawiska jak blokowanie i wiele innych zjawisk uczenia się neuronów, jest model matematyczny Harry’ego Klopfa. Model Klopfa odtwarza bardzo wiele zjawisk biologicznych, a ponadto jest prosty w implementacji.
Uogólnienie i stabilność
Reguła Hebb’a jest często uogólniana jako
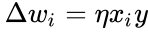
lub zmiana 
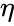
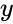
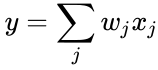
a uproszczenie z poprzedniego rozdziału przyjmuje, że zarówno współczynnik uczenia jak i wagi wejściowe wynoszą 1. Ta wersja reguły jest w oczywisty sposób niestabilna, gdyż w każdej sieci z dominującym sygnałem wagi synaptyczne będą rosły lub malały wykładniczo. Można jednak pokazać, że dla dowolnego modelu neuronu reguła Hebba jest niestabilna. Dlatego modele sieci neuronów zwykle wykorzystują inne teorie uczenia się, takie jak teoria BCM, reguła Oja lub uogólniony algorytm Hebba.
Zobacz także
- Uczenie antyhebbowskie
- Teoria BCM
- Detekcja koincydencji w neurobiologii
- Zasada Dalego
- Uogólniony algorytm Hebbiana
- Leabra
- Długotrwałeterm potentiation
- Memory
- Metaplastyczność
- Sieci neuronowe
- Reguła uczenia się Oja
- Stymulacja tetanowa
- Plastyczność zależna od czasu trwania kolców
- Hipoteza synapptotropowa
- Shouval, Harel Fizyka mózgu. Synaptic basis for Learning and Memory: A theoretical approach. The University of Texas Health Science Center at Houston. URL accessed on 2007-11-14.
Dalsza lektura
- Hebb, D.O. (1949), The organization of behavior, New York: Wiley
- Hebb, D.O. (1961). „Distinctive features of learning in the higher animal” J. F. Delafresnaye (Ed.) Brain Mechanisms and Learning, London: Oxford University Press.
- Hebb, D.O., and Penfield, W. (1940). Ludzkie zachowanie po rozległym obustronnym usunięciu z płatów czołowych. Archives of Neurology and Psychiatry 44: 421-436.
- Allport, D.A. (1985). „Distributed memory, modular systems and dysphasia” Newman, S.K. and Epstein, R. (Eds.) Current Perspectives in Dysphasia, Edinburgh: Churchill Livingstone.
- Bishop, C.M. (1995). Neural Networks for Pattern Recognition, Oxford: Oxford University Press.
- Paulsen, O., Sejnowski, T. J. (2000). Naturalne wzorce aktywności i długotrwała plastyczność synaptyczna. Current opinion in neurobiology 10 (2): 172-179.
- Overview
- Hebbian Learning tutorial (Part 1: Novelty Filtering, Part 2: PCA)
|
v-d-e
Neuroetologia |
||
|---|---|---|
| Koncepcje w neuroetologii |
Feedforward – Coincidence detector -. Umwelt – Instynkt – Detektor cech – Centralny generator wzorców (CPG) -Receptor NMDA – Hamowanie boczne – Stały wzorzec działania – Zasada Krogha – Teoria Hebrajska – Lokalizacja dźwięku |
200px
|
| Historia neuroetologii |
Theodore Holmes Bullock – Walter Heiligenberg -Niko Tinbergen- Konrad Lorenz- Eric Knudsen-Donald Griffin – Donald Kennedy – Karl von Frisch – Erich von Holst – Jörg-.Peter Ewert |
|
| Methods in Neuroethology |
Whole Cell Patch Clamp |
|
| Model Systems in Neuroethology |
Animal Echolocation – Waggle Dance- Electric Fish – Vision in toads – Neuroethology of Frog Audition – Infrared sensing in snakes |
|
Ta strona wykorzystuje treści z licencji Creative Commons pochodzące z Wikipedii (view authors).
Dodaj komentarz