” Jak duża jest komórka E. coli i jaka jest jej masa?
On 3 grudnia, 2021 by adminJak duża jest komórka E. coli i jaka jest jej masa?
Tryb czytelnika
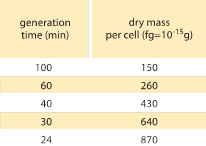
Tabela 1: Zależność między masą bakterii a czasem podziału. Sucha masa na komórkę jest podana jako funkcja czasu generacji (podwojenia). Sugeruje się, że masa wzrasta w przybliżeniu wykładniczo z szybkością wzrostu, jak pierwotnie zaobserwowali M. Schaechter et al J. Gen. Microbiol., 19:592, 1958. Sucha masa komórek została obliczona przy użyciu wartości 173 µg na jednostkę OD460 w jednym mL (BNID 106437). Użyto szczepu B/r, szczepu powszechnie stosowanego we wczesnych badaniach fizjologii bakterii. Wartości zaczerpnięte z F. C. Neidhardt, „Escherichia coli and Salmonella: Cellular and Molecular Biology”, Vol. 1., Chapter 3, ASM Press, 1996.
Rozmiar typowej bakterii takiej jak E. coli służy jako wygodna standardowa linijka do charakteryzowania skal długości w biologii molekularnej i komórkowej. Zasadą kciuka” opartą na pokoleniach pomiarów mikroskopii świetlnej i elektronowej dla wymiarów komórki E. coli jest przypisanie jej średnicy około ≈1µm, długości ≈2µm i objętości ≈1µm3 (1 fL) (BNID 101788). Kształt ten można w przybliżeniu określić jako sferocylinder, czyli cylinder z półkulistymi nasadami. Biorąc pod uwagę podaną średnicę i długość możemy obliczyć bardziej precyzyjne oszacowanie objętości ≈1,3 µm3 (dokładnie 5π/12). Różnica pomiędzy tą wartością a podaną powyżej wartością wynikającą z reguły kciuka pokazuje poziom niespójności, z którym wygodnie jest nam żyć stosując reguły kciuka. Jedną z najprostszych dróg do oszacowania masy bakterii jest wykorzystanie ≈1 µm3 objętości komórki E. coli i założenie, że ma ona taką samą gęstość jak woda. To naiwne oszacowanie skutkuje inną standardową wartością, a mianowicie, że bakteria taka jak E. coli ma masę ≈1 pg (pico=10-12). Ponieważ większość komórek składa się w około 2/3 z wody (BNID 100044, 105482), a inne składniki, jak białka, mają charakterystyczną gęstość około 1,3 razy większą od gęstości wody (BNID 101502, 104272), przeliczenie z objętości komórki na masę jest dokładne do około 10%.
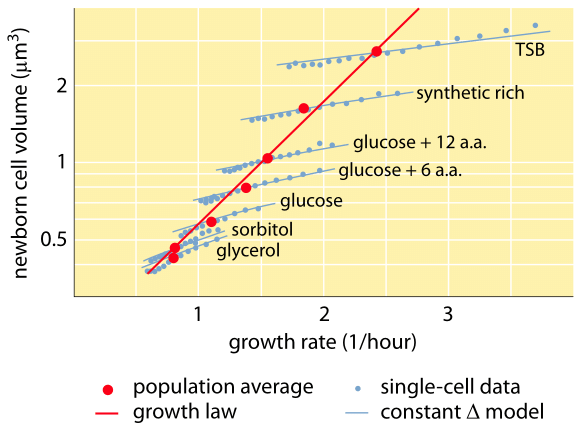
Rys. 1: Zależność między objętością komórek a tempem wzrostu. Za pomocą mikroskopii i urządzeń mikrofluidycznych można zmierzyć objętość komórki na poziomie pojedynczej komórki w różnych warunkach, potwierdzając, że średnia objętość komórki rośnie wykładniczo wraz z tempem wzrostu. Natomiast zmienność pomiędzy komórkami dla danego warunku skaluje się inaczej. Zmienność w zachowaniu pojedynczych komórek jest wykorzystywana do testowania modeli regulacji wielkości komórek. (Adaptowane z S. Taheri-Araghi et al., Curr. Biol. 25:385, 2015.)
Jeden z klasycznych wyników fizjologii bakterii podkreśla, że plastyczność właściwości komórek wynika z zależności masy komórki od tempa wzrostu. Mówiąc prościej, szybsze tempo wzrostu wiąże się z większymi komórkami. Ta obserwacja odnosi się do zmian fizjologicznych, gdzie media, które zwiększają tempo wzrostu, dają również większe komórki, jak pokazano na Rysunku 1. Zostało to również uznane za prawdziwe genetycznie, gdzie długoterminowe badania ewolucji eksperymentalnej, które doprowadziły do szybszego tempa wzrostu wykazały większe objętości komórek (BNID 110462). Takie obserwacje pomagają nam obalić mit „komórki” – gdzie ludzie, często nieświadomie, używają pomiarów dotyczących jednej komórki, aby wnioskować o innych typach komórek lub o tym samym typie komórek w innych warunkach. Klasyczne badania przeprowadzone przez Dennisa i Bremera usystematyzowały te pomiary i wykazały, że sucha masa zmienia się, jak pokazano w Tabeli 1, od średniej wartości 148 fg dla komórek dzielących się co 100 minut do 865 fg dla tych z 24-minutowym czasem podziału, co wskazuje na ponad 5-krotną różnicę w zależności od tempa wzrostu. Podobny trend zaobserwowano u innych organizmów (np. dla drożdży pączkujących, BNID 105103). Przy około 70% wody wartości te odpowiadają zakresowi od około 0,4 do 2,5 µm3 w odniesieniu do objętości. Jak można zracjonalizować większe rozmiary komórek rosnących w szybszym tempie? To pytanie jest przedmiotem debaty do dziś (Molenaar D. et al. MSB 5:323, 2009; Amir, A., Phys. Rev, Let., 112:208102, 2014). Wyjaśnienia różnią się od sugerowania, że ma on przewagę w sposobie alokacji zasobów, do twierdzenia, że jest to właściwie tylko efekt uboczny posiadania wbudowanego okresu około 60 minut od czasu, gdy komórka zdecyduje, że zgromadziła wystarczającą masę, aby rozpocząć przygotowania do podziału i do czasu, gdy zakończy replikację DNA i akt podziału. Ten z grubsza stały okres „opóźnienia” prowadzi do wykładniczej zależności średniej masy komórki od tempa wzrostu w tym toku rozumowania (Amir, A., Phys. Rev, Let., 112:208102, 2014).
Metody pomiaru objętości komórek wahają się od użycia licznika Coultera ((BNID 100004), który wnioskuje o objętości na podstawie zmian w oporze małej kryzy, gdy komórka przechodzi w niej, do bardziej bezpośrednich pomiarów przy użyciu mikroskopii fluorescencyjnej, które mierzą długości i średnice komórek w różnych warunkach (Rysunek 1 i BNID 106577, 111480). Zaskakująco, fakt, że różne laboratoria nie zawsze są zgodne co do tych samych wartości może być spowodowany różnicami w metodach kalibracji lub dokładnych szczepach i warunkach wzrostu. Bezprecedensowa zdolność do pomiaru masy komórek jest osiągana poprzez efektywne ważenie komórek na mikroskopijnym wsporniku. Jak pokazano na rysunku 2A, przepływ płynu jest wykorzystywany do wymuszania ruchu komórek tam i z powrotem w wydrążonym wsporniku. Pomiar wykorzystuje fakt, że masa komórki wpływa na częstotliwość oscylacji wspornika. Częstotliwość ta może być mierzona z niezwykłą dokładnością i wykorzystywana do wnioskowania o masie z femtogramową precyzją. Poprzez zmianę kierunku przepływu cieczy, komórka zostaje uwięziona na kilka minut lub dłużej, a tempo akumulacji jej masy jest mierzone w sposób ciągły na poziomie pojedynczej komórki. W początkowym zastosowaniu tej techniki wykazano, że pojedyncze komórki, które są większe, również szybciej gromadzą masę, rzucając światło na od dawna zadawane pytanie: czy wzrost komórek jest liniowy w czasie, czy też bardziej odpowiedni jest trend w przybliżeniu wykładniczy? Różnice mogą być niewielkie, ale dzięki tym rewolucyjnym możliwościom wyraźnie widać, że ten drugi scenariusz lepiej reprezentuje sytuację w kilku badanych typach komórek, jak pokazano na Rysunku 2B.
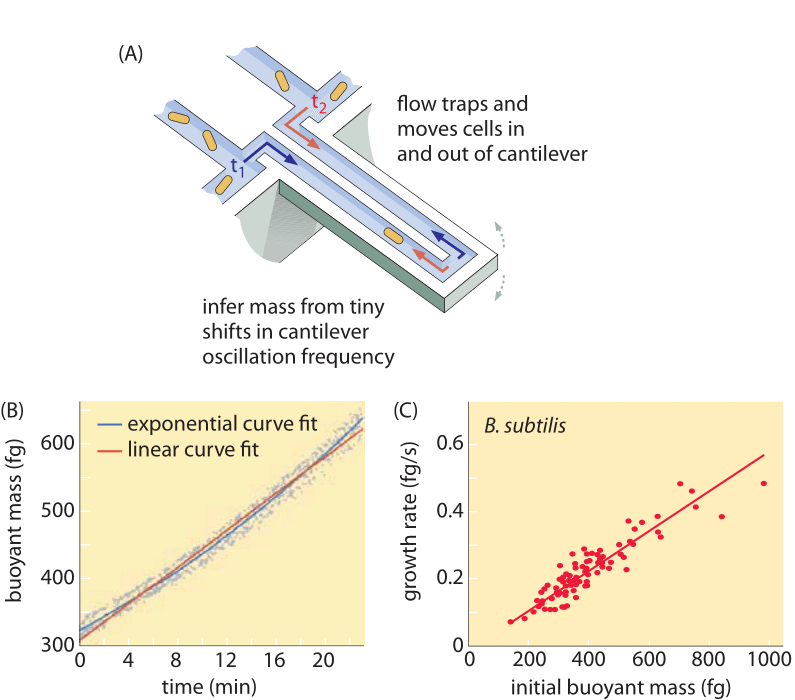
Rys. 2: Wykorzystanie masy wyporowej do pomiaru wzrostu pojedynczych komórek. (A) Mikronowy wspornik oscyluje z wysoką częstotliwością, a masa komórek może być określona na podstawie zmian w częstotliwości oscylacji. (B) Mierząc w czasie, uzyskuje się krzywą akumulacji masy pojedynczych komórek, jak pokazano na rysunku. (C) Pokazane są tu komórki B. subtilis. Porównanie przewidywań liniowego i wykładniczego modelu wzrostu pokazane jest jako najlepsze dopasowanie. Podobieństwo pokazuje jak blisko siebie są te dwa modele w zakresie zaledwie dwukrotnego wzrostu w trakcie cyklu komórkowego. Sucha masa komórki jest około 4 razy większa od masy wypornościowej. (Adaptowane z M. Godin et al., Nature Meth. 7:387, 2010.)





.
Dodaj komentarz