Funkcja Gompertza
On 12 grudnia, 2021 by adminKrzywa GompertzaEdit
Biologia populacji zajmuje się w szczególności funkcją Gompertza. Funkcja ta jest szczególnie przydatna w opisie szybkiego wzrostu pewnej populacji organizmów, a jednocześnie jest w stanie uwzględnić ostateczną asymptotę poziomą, po określeniu pojemności nośnej (plateau liczby komórek/populacji).
Modeluje się ją w następujący sposób:

gdzie:
- t jest czasem
- N0 jest początkową ilością komórek
- NI jest plateau liczby komórek/populacji
- b jest początkowym tempem wzrostu guza
Ta funkcja uwzględniająca plateau liczby komórek czyni ją użyteczną w dokładnym naśladowaniu dynamiki rzeczywistej populacji. Funkcja jest również zgodna z funkcją sigmoidalną, która jest najbardziej powszechnie akceptowaną konwencją ogólnego opisu wzrostu populacji. Ponadto, funkcja wykorzystuje początkowe tempo wzrostu, co jest powszechnie spotykane w populacjach komórek bakteryjnych i nowotworowych, które przechodzą fazę logarytmiczną i szybko rosną w liczebności. Pomimo swojej popularności, funkcja początkowego tempa wzrostu guza jest trudna do określenia, biorąc pod uwagę różne mikrokosmosy występujące u pacjenta lub różne czynniki środowiskowe w przypadku biologii populacyjnej. U pacjentów z nowotworami czynniki takie jak wiek, dieta, pochodzenie etniczne, predyspozycje genetyczne, metabolizm, styl życia i pochodzenie przerzutów odgrywają rolę w określaniu tempa wzrostu guza. Oczekuje się, że pojemność nośna również będzie się zmieniać w oparciu o te czynniki, dlatego opisywanie takich zjawisk jest trudne.
Krzywa metabolicznaEdit
Funkcja metaboliczna zajmuje się w szczególności określaniem tempa metabolizmu w organizmie. Funkcja ta może być stosowana do monitorowania komórek nowotworowych; tempo metabolizmu jest dynamiczne i jest bardzo elastyczne, dzięki czemu jest bardziej precyzyjne w szczegółowym określaniu wzrostu nowotworu. Krzywa metaboliczna bierze pod uwagę energię, jaką dostarcza organizm w utrzymaniu i tworzeniu tkanek. Ta energia może być uważana za metabolizm i podąża za określonym wzorcem w podziale komórkowym. Zachowanie energii może być wykorzystane do modelowania takiego wzrostu, niezależnie od różnej masy i czasu rozwoju. Wszystkie taksony mają podobny wzorzec wzrostu i model ten, w rezultacie, uwzględnia podziały komórkowe, podstawę rozwoju guza.

- B = energia zużywana przez organizm w spoczynku
- NC = liczba komórek w danym organizmie
- BC= tempo metabolizmu pojedynczej komórki
- NCBC= energia wymagana do utrzymania istniejącej tkanki
- EC= energia potrzebna do utworzenia nowej tkanki z pojedynczej komórki
Różnicowanie pomiędzy energią zużywaną w spoczynku a pracą w tempie metabolicznym pozwala na dokładniejsze określenie w modelu tempa wzrostu. Energia w spoczynku jest niższa od energii zużywanej do utrzymania tkanki, a razem stanowią energię potrzebną do utrzymania istniejącej tkanki. Wykorzystanie tych dwóch czynników, obok energii wymaganej do stworzenia nowej tkanki, kompleksowo odwzorowuje tempo wzrostu, a ponadto prowadzi do dokładnej reprezentacji fazy opóźnienia.
Wzrost guzówEdit
W latach 60-tych A.K. Laird po raz pierwszy z powodzeniem wykorzystał krzywą Gompertza do dopasowania danych dotyczących wzrostu guzów. W rzeczywistości, guzy są populacjami komórek rosnącymi w zamkniętej przestrzeni, gdzie dostępność składników odżywczych jest ograniczona. Oznaczając rozmiar guza jako X(t) użyteczne jest zapisanie krzywej Gompertza w następujący sposób:
X ( t ) = K exp ( log ( X ( 0 ) K ) exp ( – α t ) ) {}

gdzie:
- X(0) jest rozmiarem guza w początkowym czasie obserwacji;
- K jest nośnością, tj.tj. maksymalny rozmiar, który może być osiągnięty przy dostępnych składnikach odżywczych. W rzeczywistości jest to:

niezależnie od X(0)>0. Zauważmy, że przy braku terapii itp. zwykle jest to X(0)<K, podczas gdy w obecności terapii może to być X(0)>K;
- α jest stałą związaną ze zdolnością proliferacyjną komórek.
- log() odnosi się do logu naturalnego.
Można pokazać, że dynamiką X(t) rządzi równanie różniczkowe Gompertza:

i.e. ma postać po rozbiciu:

F(X) jest chwilową szybkością proliferacji populacji komórek, którego malejący charakter wynika z konkurencji o składniki odżywcze spowodowanej wzrostem populacji komórek, podobnie jak w przypadku logistycznego tempa wzrostu. Istnieje jednak zasadnicza różnica: w przypadku logistycznym tempo proliferacji dla małej populacji komórkowej jest skończone:

gdzie w przypadku Gompertza tempo proliferacji jest nieograniczone:

Jak zauważyli Steel i Wheldon, tempo proliferacji populacji komórkowej jest ostatecznie ograniczone przez czas podziału komórki. Może to więc świadczyć o tym, że równanie Gompertza nie jest dobre do modelowania wzrostu małych guzów. Co więcej, ostatnio zauważono, że uwzględniając interakcję z systemem immunologicznym, równanie Gompertza i inne prawa charakteryzujące się niezwiązanym F(0) wykluczałyby możliwość nadzoru immunologicznego.
Badania teoretyczne Fornalskiego i wsp. wykazały biofizyczne podstawy krzywej Gompertza dla wzrostu nowotworów, z wyjątkiem bardzo wczesnej fazy, gdzie funkcja paraboliczna jest bardziej odpowiednia. Stwierdzili również, że krzywa Gompertza opisuje najbardziej typowy przypadek wśród szerokiej rodziny funkcji dynamiki nowotworów.
Rozrost Gompertza i wzrost logistycznyEdit
Równanie różniczkowe Gompertza
jest przypadkiem granicznym uogólnionego logistycznego równania różniczkowego

(gdzie ν > 0 {{frac {1}{nu >0}

jest dodatnią liczbą rzeczywistą), ponieważ

.
W dodatku, istnieje punkt przegięcia na wykresie uogólnionej funkcji logistycznej, gdy

i jeden na wykresie funkcji Gompertza, gdy

.
Modelowanie trajektorii infekcji COVID-19Edit

Uogólniona funkcja logistyczna, zwana również krzywą wzrostu Richardsa, jest szeroko stosowana w modelowaniu trajektorii infekcji COVID-19. Trajektoria zakażenia jest dziennym szeregiem czasowym danych dotyczących skumulowanej liczby zakażonych przypadków dla danego podmiotu, takiego jak kraj, miasto, stan itp. W literaturze istnieją różne warianty re-parametryzacji: jedną z często używanych form jest
f ( t ; θ 1 , θ 2 , θ 3 , ξ ) = θ 1 1 / ξ {displaystyle f(t;θta _{1},θta _{2},θta _{3},ξ )={frac { θta _{1}}{^{1/ ξ }}}}

gdzie θ 1 , θ 2 , θ 3 {frac {theta _{1},θ _{2},θ _{3}}}.

są liczbami rzeczywistymi, a ξ {displaystyle \xi }

jest dodatnią liczbą rzeczywistą. Elastyczność krzywej f {{displaystyle f}

wynika z parametru ξ {{displaystyle \xi }
: (i) jeśli ξ = 1 {displaystyle ξ =1}

to krzywa sprowadza się do funkcji logistycznej, oraz (ii) jeśli ξ { {displaystyle ξi }
zbiega do zera, to krzywa zbiega do funkcji Gompertza. W modelowaniu epidemiologicznym, θ 1 {{displaystyle ξ _{1}}

, θ 2 {displaystyle θ _{2}}

, oraz θ 3 {displaystyle \theta _{3}}

reprezentują odpowiednio ostateczny rozmiar epidemii, współczynnik zakażenia i fazę opóźnienia. Zobacz prawy panel dla przykładowej trajektorii infekcji, gdy ( θ 1 , θ 2 , θ 3 ) {displaystyle (θtheta _{1},θ 2 , θ 3 )}

są wyznaczone przez ( 10 , 000 , 0.2 , 40 ) {displaystyle (10,000,0.2,40)}

.
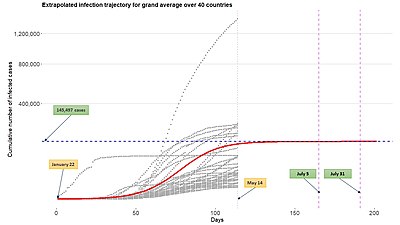
Jedną z korzyści płynących z używania funkcji wzrostu, takich jak uogólniona funkcja logistyczna w modelowaniu epidemiologicznym, jest jej stosunkowo łatwe rozszerzenie na ramy modelu wielopoziomowego poprzez użycie funkcji wzrostu do opisania trajektorii infekcji z wielu podmiotów (krajów, miast, stanów, itp.) Patrz powyższy rysunek. Takie ramy modelowania mogą być również szeroko nazywane nieliniowym modelem efektów mieszanych lub hierarchicznym modelem nieliniowym.
Prawo Gomp-ex wzrostuEdit
W oparciu o powyższe rozważania, Wheldon zaproponował model matematyczny wzrostu guza, zwany modelem Gomp-Ex, który nieznacznie modyfikuje prawo Gompertza. W modelu Gomp-Ex zakłada się, że początkowo nie ma konkurencji o zasoby, więc populacja komórek rozrasta się zgodnie z prawem wykładniczym. Istnieje jednak krytyczny próg wielkości X C {{C}}

taki, że dla X > X C {{displaystyle X>X_{C}}

. Założenie, że nie ma konkurencji o zasoby jest prawdziwe w większości scenariuszy. Mogą na nią jednak wpływać czynniki ograniczające, co wymaga stworzenia zmiennych podczynnikowych.
wzrost następuje zgodnie z prawem Gompertza:

więc:

Tutaj są pewne numeryczne oszacowania dla X C {displaystyle X_{C}}
Dodaj komentarz