Frontiers in Psychology
On 28 września, 2021 by adminCiało modzelowate (CC) jest podstawową szczeliną łączącą dwie półkule mózgowe mózgu. Dlatego też, podczas gdy termin commissurotomia nie jest niepoprawny dla opisania tak zwanego „rozdwojenia mózgu” i jest ściśle poprawny dla pierwotnych operacji chirurgicznych u ludzi, w których wszystkie ciała modzelowate zostały przecięte (Gazzaniga i in., 1967), późniejsze operacje odcięły tylko CC, wprowadzając tym samym do użycia termin „callosotomia”, który pozostaje jako bardziej odpowiedni w tych przypadkach (np. Franz i in., 1996, 2000).
Z perspektywy ewolucyjnej, CC jest niedawnym rozwojem obserwowanym tylko u ssaków łożyskowych (Katz i in., 1983; Mihrshahi, 2006). Uważa się, że funkcjonalność zapewniana przez CC jest poprzedzona, przynajmniej częściowo, przez przecinek przedni (AC), jak zaobserwowano u ssaków nie łożyskowych, takich jak marsupialce (Heath i Jones, 1971; Ashwell i in., 1996). Chociaż niektóre szczątkowe drogi AC są nadal obecne w ludzkim mózgu, ich połączenia i struktura pozostają słabo zbadane w obecnej literaturze, a nieliczne wyniki badań są raczej niespójne (Lasco i in., 2002; Patel i in., 2010; Choi i in., 2011). Na przykład, ustalenia oparte na obrazowaniu rezonansem magnetycznym (MRI) są niejednoznaczne co do tego, czy AC ma podobną wielkość u kobiet i mężczyzn (Lasco i in., 2002; Patel i in., 2010). W stosunkowo niedawnym badaniu wykorzystującym technikę in-vivo obrazowania tensora dyfuzji (DTI), która wykorzystuje dyfuzję cząsteczek wody w aksonach, Patel i wsp. (2010) przebadali osiem osób i z powodzeniem wyizolowali włókna AC u pięciu osób. U jednego z badanych stwierdzono, że duża część włókien w tylnej kończynie AC wędrowała obustronnie do okolic ciemieniowych, gdzie większość z nich kończyła się, mniejsza wiązka kończyła się w zakręcie przedśrodkowym, a jeszcze inna wiązka wchodziła do płata skroniowego. Takie różnice indywidualne stwierdzono u wszystkich uczestników, jednak większość badanych wykazywała obustronne projekcje do kory oczodołowo-czołowej, ciemieniowej i skroniowej, do których CC nie wykazuje wyraźnych projekcji. Łącznie, ustalenia te sugerują, że AC ma zarówno pierwotne jak i śladowe projekcje do obszarów neokorowych, do których CC nie rzutuje, jak sugerowali inni (Di Virgilio i in., 1999; Patel i in., 2010).
W naszym przekonaniu, AAA w dużym stopniu opiera się na sieciach neokorowych i łączności ze strukturami podkorowymi, takimi jak zwoje podstawy i wzgórze (wśród innych, nie omówionych szczegółowo w naszej propozycji) (Franz, 2012). Tak więc, wszelkie ewolucyjne zmiany dotyczące AC mogą mieć również pewną rolę w modelu AAA. Jednak znaleźliśmy niewiele literatury konkretnie zajmującej się tym możliwym powiązaniem.
Rola AC była badana w zadaniach, które prawdopodobnie będą wymagać form uwagi, takich jak te obejmujące uwagę wzrokową. Na przykład, dobrze wiadomo, że populacje acallosal były badane w kontekście zadań obejmujących integrację wzrokową (Gazzaniga i in., 1962, 1965; Sperry, 1968; Levy i in., 1972). Ostatnio Corballis (1995) starał się zająć integracją wzrokową i uwagą w celu wyjaśnienia wcześniejszych rozbieżności w badaniach nad akralgią. Corballis zaproponował dychotomiczny system uwagi wzrokowej; jedna część (obejmująca formę) jest automatyczna i zlokalizowana w każdej półkuli, a druga (obejmująca ruch i lokalizację) jest dobrowolna i podkorowa. Model AAA może bazować na tej obserwacji i odpowiedzieć na rozbieżności obserwowane w literaturze. W oparciu o zaproponowany przez nas model, na wielu poziomach uwagi istnieją drogi inne niż CC, które mogą pośredniczyć w przekazywaniu informacji między półkulami. Jedna z możliwości dotyczy obwodów zwojów podstawy, które, jak proponujemy, są potencjalnie krytyczne w pośredniczeniu w dzieleniu uwagi między półkulami (Franz, 2012). Poprzez przecięcie CC bezpośredni transfer uwagi poprzez drogi korowe jest w dużej mierze zniesiony (o ile, oczywiście, AC nie odgrywa roli rezerwowej). Jednak integracja informacji wzrokowych oparta na uwadze może być nadal pośredniczona przez drogi podkorowe i/lub AC. W ten sposób dwuczęściowy system uwagi obserwowany przez Corballisa (1995) mógłby być obecny dzięki opartemu na uwadze pośredniczeniu w integracji wzrokowej przez AC (i/lub zwoje podstawy i związane z nimi obwody: Franz, 2012).
Badania dotyczące widzenia i uwagi nie muszą być rozpatrywane całkowicie oddzielnie od systemu regulującego działanie motoryczne. W oparciu o nasze własne odkrycia (Franz, 2004; Franz i Packman, 2004), różne formy uwagi (wzrokowa i wewnętrzna-niewzrokowa) są ściśle powiązane z działaniem motorycznym, a to oznacza, że możliwy klucz do zrozumienia potencjalnej roli uwagi AC leży w związku między działaniem, widzeniem i uwagą. We wcześniejszym badaniu Franz (2004) kazał normalnym uczestnikom rysować okręgi oburęcznie pod wpływem różnych manipulacji wizualnym sprzężeniem zwrotnym lub wewnętrzną uwagą na każdą rękę. Rozmiar kół rysowanych przez rękę, która otrzymała pewną formę wewnętrznej uwagi, stał się większy w sposób podobny do efektów, które wystąpiły przy obecności wizualnego sprzężenia zwrotnego dla tej ręki (w stosunku do braku uwagi/wizualnego sprzężenia zwrotnego). Nasz model AAA implikuje, że uwaga jest kluczowym komponentem w regulacji wyjścia motorycznego (jak również wyboru planowanego działania motorycznego) i w świetle obecnych dowodów nie możemy wykluczyć, że AC odgrywa rolę w tej funkcji. Jednakże, bezpośrednie badanie roli AC w jakimkolwiek zadaniu poznawczym nie jest trywialne.
Testowanie czy AC bierze udział w funkcji poznawczej jest prawie niemożliwe w badaniach neuropsychologicznych biorąc pod uwagę, że uczestnicy z zlokalizowanym AC nie są łatwi do znalezienia (i nie wiemy o żadnym). Dlatego o wiele bardziej intuicyjne jest ocenianie neurologicznych różnic w normalnym AC i korelowanie danych neurologicznych z danymi behawioralnymi. Przedstawiona metoda traktografii (objaśniająca AC) może być stosunkowo łatwo zastosowana w dużych próbach normalnej populacji, i idealnie, może być połączona z wydajnością (opartą na zadaniach przeprowadzonych podczas oddzielnych sesji testowych z kolekcji DTI) tych samych osób przy użyciu celowo zaprojektowanych zadań poznawczych. W tym miejscu demonstrujemy, że DTI może być przeprowadzona i może przynieść użyteczne wyniki na małej próbie osób z neurologicznie normalnej populacji. Ogólna propozycja polegałaby na przeprowadzeniu podobnych metod na większej próbie i (jak zasugerowano powyżej), skorelowaniu ustaleń (przybliżonej miary gęstości AC, na przykład) ze zmiennymi w zadaniach poznawczych.
Na Rysunku 1 pokazujemy wyniki śledzenia DTI na danych uzyskanych od 10 uczestniczek otrzymanych z publicznego zbioru danych DTI na Uniwersytecie Johnsa Hopkinsa (zobacz Podziękowania dla pełnego źródła). Średnia wieku wynosiła 22 lata (SD = 2,5). Obrazy przetwarzano przy użyciu FSL Diffusion Toolbox (Smith i in., 2004; Woolrich i in., 2009; Jenkinson i in., 2012), a procedury wykorzystano w poprzednich badaniach nad innymi szlakami włóknistymi (FDT; Behrens i in., 2003a,b, 2007; Johansen-Berg i in., 2004). W skrócie, obrazy były najpierw konkatenowane do objętości 4d dla każdego uczestnika, a czaszki rozbierane przy użyciu narzędzia Brain Extraction Tool (BET; Smith, 2002; Jenkinson i in., 2005) firmy FSL. Wyekstrahowane obrazy zostały skorygowane o niewielkie ruchy głowy i prądy wirowe generowane przez zmiany kierunku pola. Dopasowywano tensory dyfuzji w celu wygenerowania obrazów przedstawiających główne kierunki dyfuzji i poziomy FA w każdym wokselu, które miały być użyte do map kolorów podczas maskowania. Bayesowska estymacja parametrów dyfuzji uzyskana przy użyciu technik próbkowania z krzyżującymi się włóknami (BEDPOSTX; „X” oznaczające krzyżujące się włókna) została przeprowadzona w celu wygenerowania rozkładu wielkości i kierunku dyfuzji uwzględniającego dwa krzyżujące się włókna, aby później przeprowadzić traktografię przez woksele dyfuzji.
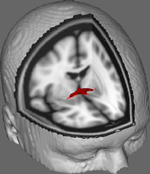
Rysunek 1. Uśrednione wyniki śledzenia przez 10 uczestników kontrolnych nałożone na standardowy obraz T1-ważony MRI z 2 mm izotropowymi wokselami dla odniesienia anatomicznego.
Uśrednione wyniki śledzenia przez wszystkich 10 uczestników kontrolnych zostały następnie nałożone na standardowy obraz T1-ważony MRI z 2 mm izotropowymi wokselami dla odniesienia anatomicznego. Średni stosunek FA w grupie wynosił 0,3146 (SD = 0,0472). Te wyniki traktografii ujawniają, co ważne, że struktura AC może być wyjaśniona, a zatem ma potencjał dla dalszych badań korelujących wyniki wydajności w zadaniach behawioralnych z wartościami FA przy użyciu większej wielkości próbki.
Podsumowując, obecnie istnieje skąpa literatura, a z dostępnej literatury żadna konkretnie nie łączy AC z procesami związanymi z modelem AAA, który jest przedmiotem naszego wcześniejszego artykułu docelowego (Franz, 2012). Stosując procedurę podobną do tej zaproponowanej w niniejszym dokumencie, możliwe jest przetestowanie większych prób normalnej populacji i skorelowanie wartości FA wyników DTI (na przykład) z konkretną zmienną zależną uzyskaną z celowo opracowanego zadania uwagi na działanie (którego procedury są w trakcie opracowywania w naszym laboratorium).
Oświadczenie o konflikcie interesów
Autorzy oświadczają, że badania zostały przeprowadzone przy braku jakichkolwiek komercyjnych lub finansowych relacji, które mogłyby być interpretowane jako potencjalny konflikt interesów.
Podziękowania
Wdzięczni jesteśmy Human Brain Project i National Research Resource Center (USA) oraz Uniwersytetowi Johna Hopkinsa za umożliwienie publicznego wykorzystania ich danych DTI. Jesteśmy również wdzięczni za hojne wsparcie grantowe z Marsden Fund of the Royal Society of New Zealand (dla Elizabeth A. Franz).
Corballis, M. (1995). Visual integration in the split brain. Neuropsychologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). The allocation of attention to learning of goal-directed actions: a cognitive neuroscience framework focusing on the basal ganglia. Front. Psychol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., and Jones, E. (1971). Interhemispheric pathways in the absence of a corpus callosum. An experimental study of commissural connexions in the marsupial phalanger. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
.
Dodaj komentarz