Superior olivary nucleus
On november 12, 2021 by admin| Hersenen: Superieur olivariscomplex | ||
|---|---|---|
 |
||
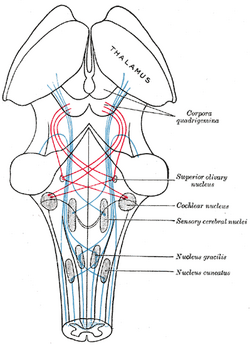
| Schema met het verloop van de vezels van de lemniscus; mediale lemniscus in blauw, laterale in rood. (De nucleus olivaris supérieur is rechts in het midden aangegeven.) | ||
| ] | ||
| Latijn | nucleus olivaris superior | |
| Gray’s | onderwerp #187 787 | |
| Deel van | ||
| Componenten | ||
| Aderslagader | ||
| Vein | ||
| BrainInfo/UW | {{BrainInfoType}}-565 | |
| MeSH | ||
Het superieure olivariumcomplex (of SOC of superior olive) is een kleine massa grijze substantie gelegen op het dorsale oppervlak van het laterale deel van het trapeziumlichaam. Klein bij de mens, maar goed ontwikkeld bij bepaalde dieren, vertoont het een soortgelijke structuur als de inferieure olivariskern, en ligt er onmiddellijk boven. Sommige van de vezels van het trapeziumlichaam eindigen door arborizing rond de cellen van deze kern, terwijl andere uit deze cellen voortkomen.
Fysiologie
De superieure olivarynucleus speelt een aantal rollen in het gehoor. De mediale superieure olijf (MSO) is een gespecialiseerde kern die wordt verondersteld om het tijdsverschil van aankomst van geluiden tussen de oren (het interaurale tijdsverschil of ITD) te meten. De ITD is een belangrijke aanwijzing voor het bepalen van het azimut van laagfrequente geluiden, d.w.z. het lokaliseren ervan in het azimutale vlak – de mate waarin zij naar links of naar rechts wijzen.
De laterale superieure olijf (LSO) wordt geacht betrokken te zijn bij het meten van het niveauverschil van geluiden tussen de oren (het interaurale niveauverschil of ILD). Het ILD is een tweede belangrijke aanwijzing bij het bepalen van het azimut van hoogfrequente geluiden.
Relatie tot het auditieve systeem
Het superieure olivariumcomplex bevindt zich in het algemeen in de hersenen, maar strekt zich bij de mens uit van de rostrale medulla tot de middenhersenen en ontvangt projecties van voornamelijk de anteroventrale cochleaire kern via de ventrale akoestische stria, hoewel de posteroventrale kern er ook projecties van heeft via de intermediaire akoestische stria. Het is op deze plaats waar de eerste belangrijke binaurale interacties optreden.
Primaire kernen
Het superieure olivarium complex is verdeeld in drie primaire kernen, de MSO, LSO, en de Mediale Nucleus van het Trapezoïd lichaam, en verscheidene kleinere periolivaire kernen. Deze drie kernen zijn het best bestudeerd, en daarom het best begrepen. Typisch worden zij beschouwd als het opgaande azimutale lokaliseringspad.
Mediale superieure olijf (MSO)
De mediale superieure olijf wordt geacht te helpen bij het lokaliseren van het azimut van een geluid, dat wil zeggen, de hoek naar links of rechts waar de geluidsbron zich bevindt. De eerste indruk zou kunnen zijn dat deze kern ook verticale informatie bevat, maar dat is niet het geval; de informatie die in de MSO wordt verwerkt, is beperkt tot het horizontale vlak. De fusiforme cellen projecteren naar niets op het niveau van de pons, en komen pas in actie bij de colliculus inferior. Er zijn alleen horizontale gegevens aanwezig, maar deze zijn wel afkomstig van twee verschillende oorbronnen, hetgeen helpt bij het lokaliseren van geluid op de azimut-as. De manier waarop de superieure colliculus dit doet is door het meten van de verschillen in tijd tussen twee oorsignalen die dezelfde stimulus registreren. De reis rond het hoofd duurt ongeveer 700 μs en er wordt aangenomen dat het mediale superieure olijf in staat is dit te detecteren. In feite is waargenomen dat mensen interaurale verschillen tot op 10 microseconden kunnen detecteren. De nucleus is tonotopisch georganiseerd, maar de azimutale receptieve veldprojectie is “hoogstwaarschijnlijk een complexe, niet-lineaire kaart.”
De projecties van de mediale superieure olijf eindigen dicht in de ipsilaterale centrale nucleus van de inferieure colliculus. De meerderheid van deze axonen worden beschouwd als “rond gevormd” of type R. Deze R axonen zijn meestal glutamatergisch en bevatten ronde synaptische vesikels en vormen asymmetrische synaptische juncties.
- Dit is de grootste van de kernen en bevat bij de mens ongeveer 15.500 neuronen.
- Elke MSO ontvangt laagfrequente bilaterale inputs van de rechter en linker AVCN’s.
- De output is naar de ipsilaterale laterale lemniscus en uiteindelijk naar de inferieure colliculus.
- De MSO reageert beter op binaurale stimuli.
- De belangrijkste functie is de detectie van verschillen in aankomsttijd van geluiden in de twee oren, wat deel uitmaakt van het lokalisatieproces (ITD).
Laterale superieure olijf (LSO)
Deze olijf heeft soortgelijke functies als de mediale superieure olijf, maar maakt gebruik van de intensiteit om de geluidsbron te lokaliseren. Dit is het deel van de hersenstam dat het hardere geluid van het linkeroor als links aanmerkt. Het laterale olijf ontvangt input van beide cochleaire kernen, van ipsilaterale AVCN sferische bushy cellen en contralaterale AVCN globulaire bushy cellen via de ipsilaterale MNTB. MNTB hoofdcellen zijn glycinerge cellen, die een bron van inhibitie vormen voor de LSO, en de basis vormen van ILD gevoeligheid. Projecties van beide cochleaire kernen zijn voornamelijk hoogfrequent, en deze frequenties worden vervolgens vertegenwoordigd door de meerderheid van de LSO neuronen (>2/3 over 2-3kHz in kat). Extra inputs zijn afkomstig van de ipsilaterale LNTB (glycinerge, zie hieronder), die remmende informatie verschaffen vanuit de ipsilaterale cochleaire kern. Een andere mogelijk remmende input is afkomstig van ipsilaterale AVCN niet-sferische cellen. Deze cellen zijn ofwel bolvormig borstelig of multipolair (stellate). Elk van deze twee inputs zou de basis kunnen vormen voor ipsilaterale inhibitie, gezien in responskaarten die de primaire excitatie flankeren, en de frequentieafstemming van de eenheid verscherpen.
De LSO projecteert bilateraal naar de centrale kern van de inferieure colliculus (ICC). Ipsilaterale projecties zijn voornamelijk inhibitoir (glycinerge), en contralateraal excitatoir. Bijkomende projectiedoelwitten zijn de dorsale en ventrale kernen van de laterale lemniscus (DNLL & VNLL). De GABA-erge projecties van de DNLL vormen de primaire bron van GABA in de auditieve hersenstam, en projecteren bilateraal naar de ICC en naar de contralaterale DNLL. Deze convergerende excitatoire en inhibitoire verbindingen kunnen de niveau-afhankelijkheid van ILD gevoeligheid in de ICC verminderen in vergelijking met de LSO.
Extra projecties vormen de Laterale Olivocochleaire Bundel (LOC), die de innerlijke haarcellen van het slakkenhuis inserveert. Aangenomen wordt dat deze projecties een lange tijdconstante hebben, en het door elk oor waargenomen geluidsniveau normaliseren om de lokalisatie van het geluid te bevorderen. Er bestaan aanzienlijke verschillen tussen de soorten: LOC projectie neuronen zijn verdeeld binnen de LSO bij knaagdieren, en omringen de LSO bij roofdieren (i.e. kat).
Medial Nucleus of Trapezoid Body (MNTB)
- De MNTB is het kleinste deel van SOC; de aanwezigheid van de MNTB in de menselijke hersenstam wordt betwijfeld.
- Elk neuron ontvangt alleen hoogfrequente input via een kelk van gehouden van de contralaterale AVCN, en zijn output projecteert naar de ipsilaterale LSO.
- Er worden twee reactietypes gevonden: een ‘chopper type’ dat lijkt op spindelcellen in de AVCN en een primair type dat lijkt op dat van Bushy Cells in de AVCN.
Periolivary Nuclei
De SOC is samengesteld uit zes tot negen periolivary kernen, afhankelijk van de geciteerde onderzoeker, gewoonlijk genoemd op basis van hun locatie ten opzichte van de primaire kernen. Deze kernen omringen elk van de primaire kernen, en dragen bij tot zowel het opgaande als het neergaande auditieve systeem. Deze kernen vormen ook de bron van de olivocochleaire bundel, die het slakkenhuis van zenuwen voorziet. Bij de cavia zijn de opgaande projecties naar de colliculi inferior hoofdzakelijk ipsilateraal (>80%), met de grootste afzonderlijke bron afkomstig van de SPON. Ook de ventrale kernen (RPO, VMPO, AVPO, & VNTB) zijn bijna volledig ipsilateraal, terwijl de overige kernen bilateraal projecteren.
| Naam | Kat | Guine Varken | Rat | Muis | |
|---|---|---|---|---|---|
| LSO | X | X | X | X | |
| MSO | X | X | X | X | |
| MNTB | X | X | X | X | |
| LNTB | X | X | “LVPO” | X | |
| ALPO | X | > | X | ||
| PVPO | X | X | |||
| PPO | X | X | “CPO” | ||
| VLPO | X | ||||
| DPO | X | X | |||
| DLPO | X | > | |||
| VTB | X | X | “MVPO” | X | |
| AVPO | X | > | |||
| VMPO | X | X | |||
| RPO | X | X | |||
| SPN | “DMPO” | X | X | X |
Ventral Nucleus of Trapezoid Body (VNTB)
- De VNTB is een kleine kern die lateraal van de MNTB ligt, en ventraal van de MSO.
- Bestaande uit een heterogene populatie van cellen, projecteert deze kern naar vele auditieve kernen, en vormt de mediale olivocochleaire bundel (MOC) die de buitenste haarcellen van het cochleair innerveert. Deze cellen bevatten elektromotiele vezels, en fungeren als mechanische versterkers / dempers binnen de cochlea.
- De kern projecteert naar beide IC, met geen cellen bilateraal projecteren.
Laterale Nucleus van het Trapeziumlichaam (LNTB)
- Locatie ventraal van de LSO
- AVCN bolvormige bossige cellen projecteren collateralen bilateraal, en bolvormige bossige cellen projecteren collateralen ipsilateraal naar LNTB neuronen.
- Cellen zijn immunoreactief voor glycine, en zijn retrogradely gelabeld na injectie van getritityleerd glycine in de LSO
- De kern projecteert naar zowel IC, met enkele cellen bilateraal projecteren, evenals de ipsilaterale LSO.
- Grote multipolaire cellen projecteren naar de cochleaire kern, maar niet de IC, in zowel kat en cavia.
- Inputs zijn vaak via end-bulbs van gehouden, produceren zeer snelle signaaltransductie.
Superior Periolivary Nucleus (SPON) (Dorsomedial Periolivary Nucleus (DMPO))
- Gelegen direct dorsaal van de MNTB
- In rat, SPON is een homogene GABA-erge kern. Deze tonotopisch georganiseerde neuronen ontvangen excitatoire inputs van octopus en multipolaire cellen in de contralaterale ventrale cochleaire kern, een glycinerge (remmende) input van de ipsilaterale MNTB, een onbekende GABA-erge (remmende) input, en projecteren naar de ipsilaterale ICC. De meeste neuronen reageren alleen op de offset van een stimulus, kunnen fase-locken op AM stimuli tot 200 Hz, en vormen mogelijk de basis voor ICC duurselectiviteit. SPON neuronen ontvangen geen dalende inputs van het IC, en projecteren niet naar het slakkenhuis of de slakkenhuiskern zoals vele periolivaire kernen doen.
- In cavia’s projecteren ronde tot ovale multipolaire cellen naar beide IC, met veel cellen die bilateraal projecteren. De meer langwerpige cellen die naar de cochleaire kern projecteren, projecteren niet naar de ICC. Er blijken twee populaties cellen te zijn, één die ipsilateraal projecteert, en één die bilateraal projecteert.
- De meeste informatie was afkomstig van knaagdier SPON, als gevolg van de prominente grootte van de kern in deze soorten, met zeer weinig studies zijn gedaan in kat DMPO, waarvan er geen waren uitgebreid.
Dorsal Periolivary Nucleus (DPO)
- Located dorsal and medial to the LSO
- Bevat zowel EE (geëxciteerd door beide oren) en E0 (geëxciteerd door het contralaterale oor alleen) eenheden.
- Neuronen zijn tonotopisch georganiseerd, en hoogfrequent.
- Kan samen met de DLPO tot één enkele kern behoren
- De kern projecteert naar beide IC, zonder bilateraal projecterende cellen.
Dorsolaterale periolivaire nucleus (DLPO)
- Ligt dorsaal en lateraal van de LSO
- Bevat zowel EE (geactiveerd door beide oren) als E0 (geactiveerd door het contralaterale oor alleen) eenheden.
- Neuronen zijn tonotopisch georganiseerd, en laagfrequent.
- Kan behoren tot een enkele kern samen met de DPO
- De kern projecteert naar beide IC, met weinig cellen die bilateraal projecteren.
Ventrolateral Periolivary Nucleus (VLPO)
- Ligt ventraal van en binnen de ventrale hillus van de LSO
- Bevat zowel EI (geëxciteerd door contralateraal en geïnhibeerd door ipsilateraal oor) als E0 (geëxciteerd door alleen het contralaterale oor) eenheden.
- Neuronen zijn tonotopisch georganiseerd, en hoog frequent.
Anterolaterale Periolivaire Nucleus (ALPO)
- De nucleus projecteert naar beide IC, met geen cellen die bilateraal projecteren.
- Grote multipolaire cellen projecteren naar de cochleaire kern, maar niet naar het IC, zowel bij de kat als bij de cavia.
Ventromediale Periolivaire Nucleus (VMPO)
- Gelegen tussen de MSO en de MNTB.
- Stuurt bilateraal projecties naar het ICC.
- De kern projecteert naar beide IC, waarbij geen cellen bilateraal projecteren.
Rostral Periolivary Nucleus (RPO) (Anterior Periolivary Nucleus (APO))
- Gelegen tussen de rostrale pool van het MSO en de VNLL
Caudale periolivaire nucleus (CPO) (posterieure periolivaire nucleus (PPO))
- Gelegen tussen de caudale pool van de MSO en de gezichtskern (7N)
Posteroventrale periolivaire nucleus (PVPO)
- De nucleus projecteert naar beide IC, zonder bilateraal projecterende cellen.
Pathofysiologie
Bij een autopsie van een 21-jarige vrouw met autisme, epilepsie en mentale retardatie werd een bijna-complete afwezigheid van de superieure olijf vastgesteld.
Zie ook
- Olivariumlichaam
- Kulesza RJ, Cytoarchitecture of the human superior olivary complex: Medial and lateral superior olive. Hearing Research 225(2007) 80-90
- 2.0 2.1 Oliver DL, et al. Axonale projecties van de laterale en mediale superieure olijf naar de inferieure colliculus van de kat: een studie met behulp van elektronenmicroscopische autoradiografie. J Comp Neurol. 1995 Sep 11;360(1):17-32 Cite error: Invalid
<ref>tag; name “R4” multiple times defined with different content - Cajal, S. R. Y. and L. Azoulay (1909). Histologie du système nerveux de l’homme et des vertébrés. Parijs, Maloine.
- 4.0 4.1 Kandel, et al Principles of Neuroscience. Vierde ed. pp 591-624. Copyright 2000, door McGraw-Hill Co. Cite error: Ongeldige
<ref>tag; naam “R2” meerdere malen gedefinieerd met verschillende inhoud - Kulesza, R. J., Jr. (2007). “Cytoarchitecture of the human superior olivary complex: medial and lateral superior olive.” Hear Res 225(1-2): 80-90.
- Tsuchitani, C. and J. C. Boudreau (1967). “Codering van stimulus frequentie en intensiteit door kat superieure olijf S-segment cellen.” J Acoust Soc Am 42(4): 794-805.
- 8.0 8.1 8.2 Cite error: Invalid
<ref>tag; name “R10” multiple times defined with different content - Wu, S. H. and J. B. Kelly (1994). “Physiological evidence for ipsilateral inhibition in the lateral superior olive: synaptic responses in mouse brain slice.” Hear Res 73(1): 57-64.
- Brownell, W. E., P. B. Manis, et al. (1979). “Ipsilateral inhibitory responses in the cat lateral superior olive.” Brain Res 177(1): 189-93.
- Darrow, K. N., S. F. Maison, et al. (2006). “Cochlear efferent feedback balanceert interaurale gevoeligheid.” Nat Neurosci 9(12): 1474-6.
- Warr, W. B. and J. J. Guinan, Jr. (1979). “Effente innervatie van het orgaan van corti: twee afzonderlijke systemen. Brain Res 173(1): 152-5.
- 13.0 13.1 Schofield, B. R. and N. B. Cant (1991). “Organisatie van het superieure olivariumcomplex bij de cavia. I. Cytoarchitecture, cytochrome oxidase histochemistry, and dendritic morphology.” J Comp Neurol 314(4): 645-70. Cite error: Invalid
<ref>tag; name “R7” multiple times defined with different content - 14.0 14.1 14.2 14.3 14.4 14.5 14.6 14.7 14.8 Illing, R. B., K. S. Kraus, et al. (2000). “Plasticiteit van het superieure olivariumcomplex. Microsc Res Tech 51(4): 364-81. Cite error: Invalid
<ref>tag; name “R5” multiple times defined with different content Cite error: Ongeldige<ref>tag; naam “R5” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R5” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R5” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R5” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R5” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R5” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R5” meerdere malen gedefinieerd met verschillende inhoud - Warr, W. B. and J. E. Beck (1996). “Multiple projections from the ventral nucleus of the trapezoid body in the rat.” Hear Res 93(1-2): 83-101.
- 16.00 16.01 16.02 16.03 16.04 16.05 16.06 16.07 16.08 16.09 16.10 Schofield, B. R. en N. B. Cant (1992). “Organisatie van het superieure olivariumcomplex bij de cavia: II. Patronen van projectie van de periolivaire kernen naar de inferieure colliculus.” J Comp Neurol 317(4): 438-55. Cite error: Ongeldige
<ref>tag; naam “R6” meerdere keren gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R6” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R6” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R6” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R6” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R6” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R6” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R6” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R6” meerdere malen gedefinieerd met verschillende inhoud Cite error: Ongeldige<ref>tag; naam “R6” meerdere malen gedefinieerd met verschillende inhoud - Smith, P. H., P. X. Joris, et al. (1993). “Projections of physiologically characterized spherical bushy cell axons from the cochlear nucleus of the cat: evidence for delay lines to the medial superior olive.” J Comp Neurol 331(2): 245-60.
- Wenthold, R. J., D. Huie, et al. (1987). “Glycine immunoreactiviteit gelokaliseerd in de cochleaire kern en superieure olivary complex. Neuroscience 22(3): 897-912.
- 19.0 19.1 Adams, J. C. (1983). “Cytology of periolivary cells and the organization of their projections in the cat. J Comp Neurol 215(3): 275-89. Cite error: Invalid
<ref>tag; name “R8” multiple times defined with different content - Friauf, E. and J. Ostwald (1988). “Divergent projections of physiologically characterized rat ventral cochlear nucleus neurons as shown by intra-axonal injection of horseradish peroxidase.” Exp Brain Res 73(2): 263-84.
- Kulesza, R. J., Jr. and A. S. Berrebi (2000). “Superior paraolivary nucleus van de rat is een GABAergic nucleus.” J Assoc Res Otolaryngol 1(4): 255-69.
- Kulesza, R. J., Jr., G. A. Spirou, et al. (2003). “Physiological response properties of neurons in the superior paraolivary nucleus of the rat.” J Neurophysiol 89(4): 2299-312.
- WHITE JS, WARR WB. The dual origins of the olivocochlear bundle in the albino rat. J. Comp. Neurol. 219:203-214, 1983.
- Guinan, J. J., Jr., S. S. Guinan, et al. (1972). “Single auditory units in the superior olivary complex. I. Respons op geluiden en classificaties gebaseerd op fysiologische eigenschappen.” Int J Neurosci 4: 101-20.
- Tsuchitani, C. (1977). “Functional organization of lateral cell groups of cat superior olivary complex.” J Neurophysiol 40(2): 296-318.
- Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996). Embryologische oorsprong voor autisme: ontwikkelingsanomalieën van de hersenzenuw motor kernen. J Comp Neurol 370 (2): 247-61.
- MedEd at Loyola Neuro/frames/nlBSs/nl27fr.htm
- Image at med.utah.edu (pick 9b. Pons)
Dit artikel was oorspronkelijk gebaseerd op een item uit een publiek domein editie van Gray’s Anatomy. Als zodanig kan een deel van de informatie hierin verouderd zijn. Indien dit het geval is, gelieve dan het artikel aan te passen, en voel je vrij om deze mededeling te verwijderen als ze niet langer relevant is.
|
v-d-e
Hersenen: rhombencephalon (achterhersenen) |
||||||
|---|---|---|---|---|---|---|
| Metencephalon/ pons |
|
Dorsaal/
(tegmentum) |
Ventraal/
(basis) |
Raphe/reticular
|
||
| Myelencephalon/ medulla |
|
Dorsal
|
Ventraal
|
Raphe/reticular
|
||
|
v-d-e
Zintuiglijk systeem: Auditieve en Vestibulaire systemen (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Buitenoor |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Earlobe) – Gehoorgang – Gehoorspieren Eardrum (Umbo, Pars flaccida) |
|||||||||||
| Middenoor |
|
|||||||||||
| Binnenoor/ (membraanhoudend labyrint, benig labyrint) |
|
|||||||||||
| {| class=”navbox collapsible nowraplinks” style=”margin:auto;” | ||||||||||||
|
—
|
||||||||||||
|
||||||||||||
|}
Deze pagina maakt gebruik van Creative Commons-gelicenseerde inhoud van Wikipedia (bekijk auteurs).
Geef een antwoord