Structuur en functie van bacteriële cellen
On oktober 10, 2021 by adminStructuur en functie van bacteriële cellen (bladzijde 5)
(Dit hoofdstuk heeft 10 bladzijden)
© Kenneth Todar, PhD
Celwand
De celwanden van bacteriën verdienen om verschillende redenen speciale aandacht:
1. Ze zijn een essentiële structuur voor levensvatbaarheid, zoals hierboven beschreven.
2. Ze zijn samengesteld uit unieke componenten die nergens anders in de natuur voorkomen.
3. Ze zijn een van de belangrijkste plaatsen voor aanvallen door antibiotica.
4. Ze leveren liganden voor binding en receptoren voor geneesmiddelen of virussen.
5. Ze veroorzaken ziektesymptomen bij dieren.
6. Ze zorgen voor immunologisch onderscheid en immunologische variatie tussen bacteriestammen.
De meeste procaryoten hebben een stijve celwand.De celwand is een essentiële structuur die de celprotoplast beschermt tegen mechanische schade en tegen osmotische breuk of lysis.Procaryoten leven gewoonlijk in een relatief verdunde omgeving, zodat de accumulatie van opgeloste stoffen binnen het cytoplasma van de procaryote cel veel groter is dan de totale absolute concentratie in het buitenmilieu. Zo kan de osmotische druk tegen de binnenzijde van het plasmamembraan het equivalent van 10-25atm bedragen. Aangezien het membraan een tere, plastische structuur is, moet het worden ingeperkt door een buitenwand van poreus, stijf materiaal met een hoge treksterkte. Een dergelijk materiaal is mureïne, de alomtegenwoordige component van bacteriële celwanden.
Mureïne is een uniek type van peptidoglycaan, een polymeer van disachariden (glycaan), vernet door korte ketens van aminozuren (peptide). Er bestaan vele soorten peptidoglycaan. Alle bacteriële peptidoglycanen bevatten N-acetylmuraminezuur, dat het bepalende bestanddeel van mureïne is. De celwanden van Archaea kunnen bestaan uit eiwitten, polysacchariden of peptidoglycaanachtige moleculen, maar bevatten nooit mureïne. Dit kenmerk onderscheidt de Bacteria van de Archaea.
In de Gram-positieve Bacteria (die welke de purperen kristalviolet kleurstof vasthouden wanneer zij aan de Gram-kleuringprocedure worden onderworpen), bestaat de celwand uit verscheidene lagen van peptidoglycan. Loodrecht op de peptidoglycanlagen staat een groep moleculen, teichoëzuren genaamd, die uniek zijn voor de Gram-positieve celwand (figuur 14). 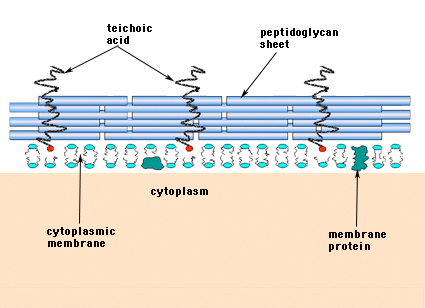
Figuur 14. Structuur van de Gram-positieve bacteriële celwand. De wand is relatief dik en bestaat uit vele lagen peptidoglycaan, afgewisseld met teichoëzuren die loodrecht op de peptidoglycaanvellen staan.
Bij de Gram-negatieve bacteriën (die het kristalviolet niet vasthouden) bestaat de celwand uit een enkele laag peptidoglycanen, omgeven door een membraanachtige structuur die het buitenmembraan wordt genoemd.Het buitenmembraan van Gram-negatieve bacteriën bevat onveranderlijk een eenduidig bestanddeel, lipopolysaccharide (LPS of endotoxine), dat giftig is voor dieren. Bij Gram-negatieve bacteriën wordt het buitenmembraan gewoonlijk beschouwd als een deel van de celwand (Figuur15).

Figuur15. Structuur van de Gram-negatieve celwand. De wand is relatief dun en bevat veel minder peptidoglycaan dan de Gram-positieve wand. Ook teichoëzuren zijn afwezig. De Gram-negatieve celwand bestaat echter uit een buitenmembraan dat zich buiten de peptidoglycaanlaag bevindt. Het buitenmembraan is verbonden met de peptidoglycaanplaat door een unieke groep lipoproteïnemoleculen.
In de Gram-positieve bacteriën is de celwand dik (15-80 nanometer), bestaande uit verschillende lagen peptidoglycaan. Bij de Gram-negatieve bacteriën is de celwand betrekkelijk dun (10 nanometer) en bestaat uit een enkele laag peptidoglycanen, omgeven door een buitenmembraan.
De structuur en rangschikking van het peptidoglycaan in E.coli is representatief voor alle Enterobacteriaceae, alsmede voor vele andere Gram-negatieve bacteriën. De glycaanruggengraat bestaat uit afwisselende moleculen van N-acetylglucosamine (G) en N-acetylmuraminezuur (M), verbonden door een bèta-1,4-glycosidebinding. De 3-koolstof van N-acetylmuraminezuur (M) is gesubstitueerd met een lactylethergroep afkomstig van pyruvaat. De lactylether verbindt de glycaanruggengraat met een peptidezijketen die L-alanine (L-ala), D-glutamaat (D-glu), diaminopimelzuur (DAP) en D-alanine (D-ala) bevat. MurNAc is uniek voor bacteriële celwanden, evenalsD-glu,DAP en D-ala. De muraminezuureenheid van E. coli wordt getoond inFiguur16.

Figuur 16. De structuur van de muraminezuur-subeenheid van het peptidoglycaan van Escherichia coli. Dit is het type mureïne dat in de meeste Gram-negatieve bacteriën wordt aangetroffen. De glycaanruggengraat is een repeterend polymeer van twee aminosuikers, N-acetylglucosamine (G) en N-acetylmuraminezuur (M). Aan het N-acetylmuraminezuur is een tetrapeptide gehecht bestaande uit L-ala-D-glu-DAP-D-ala. b. Verkorte structuur van de muraminezuursubeenheid. c. Nabijgelegen tetrapeptide zijketens kunnen met elkaar verbonden zijn door een interpeptidebinding tussen DAP op de ene keten en D-ala op de andere. d. De polymere vorm van het molecuul.
Strengen van mureïne worden in het periplasma samengesteld uit ongeveer 10 subeenheden van mureïnezuur. Vervolgens worden de strengen met elkaar verbonden tot een ononderbroken glycaanmolecuul dat de cel omvat.Waar hun nabijheid dit toelaat, kunnen de tetrapeptideketens die vanaf de glycaanruggengraat uitsteken, worden gekruist door een interpeptidebinding tussen een vrije aminogroep op DAP en een vrije carboxylgroep op een nabijgelegen D-ala.De assemblage van peptidoglycaan aan de buitenkant van het plasmamembraan wordt bemiddeld door een groep periplasmatische enzymen, namelijk transglycosylasen, transpeptidasen carboxypeptidasen. Het werkingsmechanisme van penicilline en verwante bèta-lactamantibiotica is het blokkeren van de transpeptidase- en carboxypeptidase-enzymen tijdens hun assemblage van de mureïnecelwand. Vandaar dat men zegt dat de bètalactamantibiotica de celwandsynthese in de bacteriën “blokkeren”.
De glycaanruggengraat van het peptidoglycaanmolecuul kan worden gesplitst door een enzym dat lysozym heet en aanwezig is in het dierlijk serum, weefsels en afscheidingen, en in het fagocyterende lysosoom. De functie van lysozym is het lyseren van bacteriële cellen als een constitutieve verdediging tegen bacteriële ziekteverwekkers. Sommige Gram-positieve bacteriën zijn zeer gevoelig voor lysozym en het enzym is vrij actief bij lage concentraties. Lachrymalsecreet (tranen) kan worden verdund tot 1:40.000 en behoudt het vermogen om bepaalde bacteriële cellen te lyseren. Gram-negatieve bacteriën zijn minder kwetsbaar voor lysozym omdat hun peptidoglycaan wordt afgeschermd door het buitenmembraan. De precieze plaats van de lysozym-splitsing is de bèta-1,4-binding tussen N-acetylmuraminezuur (M) en N-acetylglucosamine (G), zodat de in figuur 16(a) afgebeelde subeenheid van turaminezuur het resultaat is van de werking van het lysozym op bacterieel peptidoglycaan.
In Gram-positieve bacteriën zijn er talrijke verschillende peptidoglycans met verschillende peptidische rangschikkingen. De best bestudeerde is die van Staphylococcus aureus die in figuur 17 hieronder is afgebeeld. In plaats van DAP (in E. coli) is er het diaminozuur, L-lysine (L-lys), en in plaats van de interpeptidebinding (in Gram-negatieven) is er een interpeptidebrug van aminozuren die een vrije aminogroep op lysine verbindt met een vrije carboxylgroep op D-ala van een nabijgelegen tetrapeptidezijketen. Deze opstelling maakt kennelijk een frequentere kruisverbinding tussen nabijgelegen tetrapeptidezijketens mogelijk. In S. aureus is de interpeptidebrug een peptide dat bestaat uit 5 glycinemoleculen (een pentaglycinebrug genoemd). De vorming van de interpeptidebrug in Gram-positief mureïne wordt op dezelfde wijze geremd door bèta-lactamantibiotica als de interpeptidebrug in Gram-negatief mureïne. Gram-positieve bacteriën zijn gevoeliger voor penicilline dan Gram-negatieve bacteriën omdat het peptidoglycaan niet door een buitenmembraan wordt beschermd en het een overvloediger molecuul is. In Gram-positieve bacteriën kunnen peptidoglycanen verschillen in het aminozuur op de plaats van DAP of L-lys op positie 3 van het tetrapeptide, en in de precieze samenstelling van de interpeptidebrug. Bij Gram-positieve bacteriën komen ten minste acht verschillende typen peptidoglycanen voor.
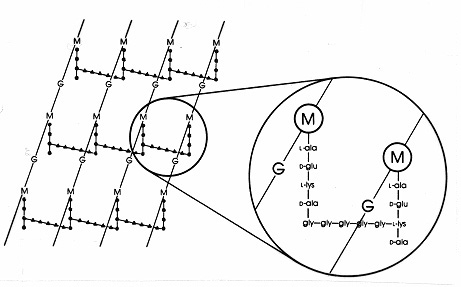
Figuur 17. Schematische weergave van de peptidoglycaanplaat van Staphylococcus aureus. G = N-acetyl-glucosamine; M = N-acetyl-muraminezuur; L-ala = L-alanine; D-ala = D-alanine; D-glu = D-glutaminezuur; L-lys = L-lysine. Dit is één type mureïne dat wordt aangetroffen in Gram-positieve bacteriën. Vergeleken met het E. coli peptidoglycan (Figuur 7) is er L-lys in plaats van DAP (diaminopimelzuur) in de tetrapeptide. De vrije aminogroep van L-lys wordt gesubstitueerd door een glycinepentapeptide (gly-gly-gly-gly-gly-) dat dan een interpeptidebrug wordt die een verbinding vormt met een carboxylgroep van D-ala in een aangrenzende tetrapeptidezijketen. Gram-positieve peptidoglycanen verschillen van soort tot soort, vooral wat betreft de aminozuren op de derde positie van de tetrapeptideketen en in de aminozuursamenstelling van de interpeptidebrug.
Gram-negatieve bacteriën kunnen in hun celwanden een enkele monomoleculaire laag mureïne bevatten, terwijl Gram-positieve bacteriën geacht worden meerdere lagen of “wikkels” van peptidoglycan te hebben. Nauw verbonden met de lagen peptidoglycaan in Gram-positieve bacteriën is een groep moleculen die teichoëzuren worden genoemd. Teichoëzuren zijn lineaire polymeren van polyglycerol of polyribitol, gesubstitueerd met fosfaten en een paar aminozuren en suikers. De teichoëzuurpolymeren zijn soms verankerd in het plasmamembraan (lipoteichoëzuur, LTA genoemd) en lijken haaks op de lagen peptidoglycaan naar buiten gericht te zijn. De functies van lipoteichonzuur zijn niet bekend. Zij zijn essentieel voor de levensvatbaarheid van Gram-positieve bacteriën in de natuur. Eén idee is dat zij een kanaal vormen van regelmatig georiënteerde negatieve ladingen om positief geladen stoffen door het gecompliceerde peptidoglycan netwerk te voeren. Een andere theorie is dat teichoïczuren op de een of andere manier betrokken zijn bij de regulering en assemblage van muraminezuureenheden aan de buitenkant van het plasmamembraan. Er zijn gevallen, met name bij de streptokokken, waarin teichoïnezuren betrokken zijn bij de hechting van de bacteriën aan weefseloppervlakken.
Hoofdstuk vervolgd
Vorige pagina
Geef een antwoord