N-gekoppelde glycosylering
On december 18, 2021 by admin
De biosynthese van N-gekoppelde glycanen verloopt via 3 grote stappen:
- Synthese van dolichol-gekoppelde precursor oligosaccharide
- En bloc overdracht van precursor oligosaccharide aan eiwit
- Verwerking van de oligosaccharide
Synthese, en bloc overdracht en initiële trimmen van precursor oligosaccharide gebeurt in het endoplasmatisch reticulum (ER). De daaropvolgende verwerking en modificatie van de oligosaccharideketen vindt plaats in het Golgi-apparaat.
De synthese van glycoproteïnen is dus ruimtelijk gescheiden in verschillende cellulaire compartimenten. Daarom hangt het type N-glycan dat wordt gesynthetiseerd af van zijn toegankelijkheid voor de verschillende enzymen die in deze cellulaire compartimenten aanwezig zijn.
Ondanks de diversiteit worden alle N-glycanen gesynthetiseerd via een gemeenschappelijke route met een gemeenschappelijke kernglycaanstructuur.De kernglycaanstructuur bestaat in wezen uit twee N-acetylglucosaminen en drie mannoseresiduen. Dit kernglycan wordt vervolgens verder uitgewerkt en gemodificeerd, wat resulteert in een uiteenlopende reeks N-glycanstructuren.
Synthese van precursor oligosaccharideEdit
Het proces van N-gekoppelde glycosylering begint met de vorming van dolichol-gekoppelde GlcNAc-suikers. Dolichol is een lipidemolecuul dat bestaat uit zich herhalende isopreeneenheden. Dit molecuul zit vast aan het membraan van de ER. Suikermoleculen zijn aan het dolichol gehecht door middel van een pyrofosfaatkoppeling (één fosfaat was oorspronkelijk aan dolichol gekoppeld, en het tweede fosfaat kwam van het nucleotidesuiker). De oligosaccharideketen wordt vervolgens uitgebreid door toevoeging van verschillende suikermoleculen op een stapsgewijze manier om een precursor oligosaccharide te vormen.
De assemblage van deze precursor oligosaccharide gebeurt in twee fasen: Fase I en II. Fase I vindt plaats aan de cytoplasmatische zijde van de ER en fase II aan de luminale zijde van de ER.
De precursormolecule, klaar om te worden overgedragen aan een eiwit, bestaat uit 2 GlcNAc, 9 mannose- en 3 glucosemoleculen.

|
|
| Fase II | |
is de mannoseresidu-donor (vorming: Dol-P + GDP-Man → Dol-P-Man + GDP) en Dol-P-Gluc is de glucoseresidu-donor (vorming: Dol-P + UDP-Glc → Dol-P-Glc + UDP).
|
|
Overdracht van glycan naar eiwitEdit
Als de precursor oligosaccharide is gevormd, wordt de voltooide glycan vervolgens overgebracht naar het ontluikende polypeptide in het lumen van het ER-membraan. Deze reactie wordt aangedreven door de energie die vrijkomt bij de splitsing van de pyrofosfaatbinding tussen het dolichol-glycaanmolecuul.Er zijn drie voorwaarden waaraan moet worden voldaan voordat een glycaan wordt overgebracht naar een nascent polypeptide:
- Asparagine moet zich bevinden in een specifieke consensussequentie in de primaire structuur (Asn-X-Ser of Asn-X-Thr of in zeldzame gevallen Asn-X-Cys).
- Asparagine moet zich op de juiste plaats in de driedimensionale structuur van het eiwit bevinden (suikers zijn polaire moleculen en moeten dus aan asparagine aan het oppervlak van het eiwit worden gehecht en niet in het eiwit worden begraven)
- Asparagine moet zich aan de luminale zijde van het endoplasmatisch reticulum bevinden om N-gekoppelde glycosylering in gang te kunnen zetten. De doelresiduen worden gevonden in secretorische proteïnen of in de naar het lumen gerichte regio’s van transmembraaneiwitten.
Oligosaccharyltransferase is het enzym dat verantwoordelijk is voor de herkenning van de consensussequentie en de overdracht van het precursorglycan naar een polypeptide-acceptor die in het endoplasmatisch reticulum lumen wordt vertaald. N-gekoppelde glycosylering is derhalve een co-translationele gebeurtenis
Verwerking van glycanEdit

N-glycanverwerking vindt plaats in het endoplasmatisch reticulum en het Golgi-lichaam. Het eerste trimmen van het precursormolecuul vindt plaats in het ER en de daaropvolgende verwerking in het Golgi.
Bij het overbrengen van het voltooide glycan op het nascente polypeptide worden twee glucose-residuen uit de structuur verwijderd. Enzymen die glycosidases worden genoemd, verwijderen sommige suikerresiduen. Deze enzymen kunnen glycosidebindingen verbreken door gebruik te maken van een watermolecuul. Deze enzymen zijn exoglycosidases, aangezien zij alleen werken op monosaccharideresiduen die zich aan het niet-reducerende uiteinde van het glycan bevinden. Aangenomen wordt dat deze eerste trimstap fungeert als een kwaliteitscontrolestap in de ER om de eiwitvouwing te controleren.
Zodra het eiwit correct is gevouwen, worden twee glucosideresiduen verwijderd door glucosidase I en II. De verwijdering van het laatste derde glucose-residu geeft aan dat het glycoproteïne klaar is voor doorvoer van het ER naar de cis-Golgi. ER mannosidase katalyseert de verwijdering van dit laatste glucose. Als het eiwit echter niet goed gevouwen is, worden de glucoseresiduen niet verwijderd en kan het glycoproteïne het endoplasmatisch reticulum niet verlaten. Een chaperon-eiwit (calnexine/calreticulin) bindt zich aan het ongevouwen of gedeeltelijk gevouwen eiwit om te helpen bij het vouwen van het eiwit.
De volgende stap bestaat uit verdere toevoeging en verwijdering van suikerresiduen in het cis-Golgi. Deze modificaties worden gekatalyseerd door respectievelijk glycosyltransferasen en glycosidasen. In de cis-Golgi verwijdert een reeks mannosidases enkele of alle van de vier mannoseresiduen in α-1,2-koppelingen. Terwijl in het mediale gedeelte van de Golgi, glycosyltransferases suikerresiduen toevoegen aan de glycaankernstructuur, waardoor de drie belangrijkste typen glycanen ontstaan: hoge mannose, hybride en complexe glycanen.
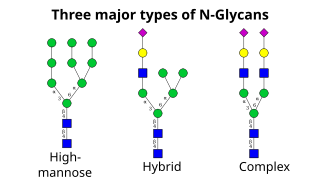
- Hoog-mannose is in wezen niet meer dan twee N-acetylglucosaminen met veel mannoseresiduen, vaak bijna evenveel als in de voorloper-oligosachariden voordat het aan het eiwit wordt gehecht.
- Complexe oligosacchariden worden zo genoemd omdat zij bijna elk aantal van de andere soorten sacchariden kunnen bevatten, waaronder meer dan de oorspronkelijke twee N-acetylglucosamines.
- Hybride oligosacchariden bevatten een mannose residu aan de ene kant van de tak, terwijl aan de andere kant een N-acetylglucosamine een complexe tak initieert.
De volgorde van toevoeging van suikers aan de groeiende glycaanketens wordt bepaald door de substraatspecificiteiten van de enzymen en hun toegang tot het substraat terwijl zij zich door de secretorische route bewegen. De organisatie van deze machinerie binnen een cel speelt dus een belangrijke rol bij het bepalen welke glycanen worden gemaakt.
Enzymen in de GolgiEdit
Golgi-enzymen spelen een sleutelrol bij het bepalen van de synthese van de verschillende soorten glycanen. De volgorde van de werking van de enzymen wordt weerspiegeld in hun positie in de Golgistapel:
| Enzymen | Locatie binnen Golgi |
|---|---|
| Mannosidase I | cis-Golgi |
| GlcNAc-transferases | mediaal Golgi |
| Galactosyltransferase en Sialyltransferase | trans-Golgi |
In archaea en prokaryotenEdit
Gelijkaardige N-glycan biosynthese paden zijn gevonden in prokaryoten en Archaea. In vergelijking met eukaryoten lijkt de uiteindelijke glycaanstructuur in eubacteriën en archaea echter niet veel te verschillen van de oorspronkelijke precursor die in het endoplasmatisch reticulum wordt gemaakt. Bij eukaryoten wordt de oorspronkelijke precursor oligosaccharide onderweg naar het celoppervlak ingrijpend gemodificeerd.
Geef een antwoord