Haaienweek: Wat at Megalodon?
On december 13, 2021 by adminGast Auteur: Jack Cooper, MSc
UoB Graduate / PhD Student, University of Swansea
Zoals we tot nu toe hebben gezien, zijn er veranderingen en updates geweest in de wetenschappelijke consensus over Megalodon’s taxonomische toewijzing en maximale lichaamsgrootte. Over zijn lievelingssnack is men het in de wetenschappelijke gemeenschap echter wel eens. Nog een zeldzaamheid, het is iets dat meestal met vrij grote nauwkeurigheid wordt afgebeeld in fictie. Sta mij toe een haai voor te stellen die walvissen at – echte walvissen!
Veel van de huidige grote macropredatoire haaien staan bekend om het eten van zeezoogdieren, vooral onze oude vriend de grote witte haai. Sommige van de meest iconische foto’s van deze haai laten zien hoe hij zich uit het water slingert met zijn laatste slachtoffer tussen zijn kaken (Fig. 1). Jonge witte haaien eten echter niet van meet af aan zeehonden. In plaats daarvan beginnen ze met het eten van vis en kleinere haaien. Als we nog eens kijken naar de Gatun formatie in Panama, vinden we veel fossiele haaien, roggen en otolieten (structuren van de binnenoren van vissen), evenals een overvloed aan ongewervelde dieren zoals weekdieren. Iets wat we echter niet vinden, zijn fossiele zeezoogdieren. Dit lijkt erop te wijzen dat Megalodon jongen ook vis en kleinere haaien aten in plaats van onmiddellijk achter zeezoogdieren aan te gaan. Dit wordt een ontogenetische verschuiving in het dieet genoemd, wat betekent dat de haai zijn voorkeursvoedselbron verandert naarmate hij groeit. Door groter voedsel te eten, en waarschijnlijk veel voedsel, toen hij volwassen werd, kon Megalodon de stofwisseling op peil houden die zo groot is. Hij kon dit doen door niet alleen walvissen te eten, maar zo ongeveer alles wat hij tegenkwam.

Gezien de hoeveelheid voedsel die Megalodon moet hebben gegeten om zijn enorme omvang te rechtvaardigen, was het vrijwel zeker een opportunistische voedseleter. Het fossielenbestand is hier onze beste gids. Dit is te danken aan de aanwezigheid van sporenfossielen, dat zijn geologische verslagen van biologische activiteit. Dit kan gaan van voetafdrukken van dinosaurussen tot krassen op botten die erop wijzen dat iets het dier heeft gebeten waartoe dat bot behoorde. En Megalodon liet een paar van deze sporenfossielen achter, die belangrijke bewijzen leveren van zijn moorden.
Er zijn verschillende fossielen gevonden van verschillende walvisachtigen met opvallende bijtsporen erop. Een goed geciteerd voorbeeld komt uit de Yorktown formatie in Maryland, waar verschillende walvisbotten bijtsporen vertoonden die duidelijk waren gemaakt door grote gekartelde tanden. Soms worden zelfs Megalodon-tanden gevonden die direct met deze botten zijn verbonden. Haaien verliezen en vervangen hun tanden duizenden keren in hun leven, en verliezen dus vaak tanden wanneer zij een gewelddadige moord begaan. Dit is een belangrijke reden waarom haaientanden zulke veel voorkomende fossielen zijn. De aanwezigheid van de haaientanden vlak naast een bekrast walvisbot wijst erop dat er werd gevoed. Maar wat paleontologen altijd moeten overwegen als deze fossielen worden gevonden, is: was dit werkelijk predatie? Of was het misschien aas-eten?
Aas-eten is een veel voorkomend voedingsgedrag bij moderne macropredatoire haaien zoals de grote witte haai (Fig. 2) en de tijgerhaai. Als deze haaien dode walvissen tegenkomen die aan het oppervlak drijven, is het een buffet van alles wat je op kunt eten. Het belangrijkste is dat het aas eten geen energie kost, iets wat verspild kan worden aan een mislukte predatiepoging. Aangezien de Megalodon zijn enorme omvang moest kunnen behouden, zou het niet erg logisch zijn om een gratis maaltijd aan zich voorbij te laten gaan als hij een dode walvis tegenkwam. Als we kijken naar hoe grote moderne haaien zich voeden, zijn de meesten van ons bereid te wedden dat Megalodon waarschijnlijk wel aasde als hij de kans kreeg.

Dit gezegd hebbende, is er één bijzonder opmerkelijk fossiel dat Megalodon betrekt bij een poging tot predatie. En, wat cruciaal is, we weten dit vooral omdat de poging mislukte. In een artikel dat in 2010 werd gepubliceerd, werd een gedeeltelijk Plioceen ribfossiel beschreven van een walvisachtige uit de Yorktown-formatie in Noord-Carolina (jawel, twee Yorktown-formaties). Ook dit fossiel vertoonde beschadigingen die leken op bijtwonden, maar opvallend genoeg bleek er een gevlochten botbedekking over deze wonden te zitten. De auteurs interpreteerden dit als een teken van een infectie die was opgetreden terwijl het dier aan het herstellen was van het bottrauma veroorzaakt door die bijtwonden. Deze sporen kwamen goed overeen met een gekartelde beet, wat suggereert dat de aanvaller gekartelde tanden had – net als Megalodon. De studie suggereerde daarom dat een Megalodon, of een andere grote haai, het dier had aangevallen waarvan het fossiel afkomstig was, maar dat het slachtoffer de poging had overleefd om zes weken later te sterven aan een infectie als gevolg van zijn verwonding.
Nog recenter werden de allereerste fossiele bijtsporen op het zuidelijk halfrond die aan Megalodon werden toegeschreven, gevonden in de Pisco-formatie van Peru. Misschien was dit wel de bedoeling, want dit is dezelfde plaats waar Gordon Hubbell zijn uitzonderlijke Carcharodon fossiel vond (besproken in Q1). Het gebeten materiaal dat hier werd gevonden omvatte schedel- en ribresten, wat suggereert dat de haai zijn doelwit van opzij had gebeten. Het belangrijkste is dat de resten voor het eerst tot een afzonderlijke soort werden geïdentificeerd – die soort is Piscobalaena nana, een kleine baleinwalvis. Net als bij de andere sporenfossielen kwamen de bijtsporen goed overeen met grote gekartelde tanden – een goed argument om Megalodon van de aanval te beschuldigen (Fig. 3). Het bleek dus dat Megalodon ook in staat was kleinere walvissen aan te vallen (of het nu om predatie of aasetende dieren ging), wat sterk overeenkomt met suggesties dat Megalodon het waarschijnlijk ook gemunt had op dolfijnen, zeeschildpadden en andere kleinere prooien.

Hoewel predatiestrategieën moeilijk te interpreteren zijn aan de hand van fossielen, kunnen moderne haaien ons wel een idee geven van hoe Megalodon jaagde. Zijn bijtkracht zou uitzonderlijk krachtig zijn geweest om walvisbotten te beschadigen. Volgens een computermodel van de schedel van een witte haai heeft deze haai een bijtkracht van meer dan 1,8 ton, waarmee hij een van de sterkste bijtkrachten van alle levende dieren heeft. In het verlengde hiervan suggereert deze studie dat de bijtkracht van Megalodon nog groter was, met een schatting van een buitengewone 108.514-182.201 N.
Wetenschappers hebben ook pogingen gedaan om de zwemsnelheid van Megalodon te berekenen, wat ons een indicatie kan geven van hoe hij jaagde. Haaien kunnen een burstsnelheid hebben, dat is een snelle zwemstoot, die typisch wordt gebruikt om een prooi te grijpen. Maar hun gebruikelijke zwemsnelheid is wat wij noemen een constante zwemsnelheid. Dit varieert van soort tot soort, maar een studie onder leiding van David Jacoby combineerde gegevens over 26 soorten uit 64 eerdere studies om een model te ontwikkelen dat een schalingsrelatie voorspelt tussen de snelheid waarmee de haai zwemt en het metabolisme en de lichaamsmassa van de haai. Hun model waarin het metabolisme is verwerkt, geeft een schalingsexponent van 0,173, samengevat in de volgende vergelijking:
Zwemsnelheid (ms ¯¹) ∝ Massa (kg)^0.173
Wanneer zij de gegevens corrigeerden voor fylogenie, stelden zij echter vast dat de minimale zwemsnelheid met de lichaamsmassa een iets lagere exponent van 0,15 had. De bovenstaande vergelijking veronderstelt bovendien een constante van 1 om overeen te komen met de verhoudingen, maar uit de gegevens bleek dat deze constante in werkelijkheid 0,266 was. Daarom construeerden zij de vermogensvergelijking:
Zwemsnelheid (ms ¯¹) = 0,266 Massa (kg) ^0,15
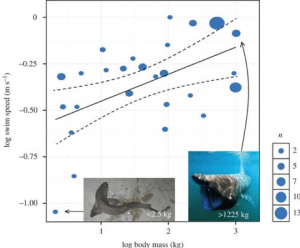
Hieruit kunnen zij de volgehouden zwemsnelheid van Megalodon berekenen, afhankelijk van zijn massa. Bijvoorbeeld, de ~48.000 kg Megalodon berekend als de massa van een 15,9 m lange haai in het werk van Gottfried, zou een haai opleveren die continu zwemt met een snelheid van 1,34 m/s – ruwweg 4,8 km/uur. Als we dit model zouden toepassen op een Megalodon van 52.000 kg, zou de zwemsnelheid 1,36 m/s of 4,9 km/uur bedragen. Als we het echt extreem zouden aanpakken en een haai van 100.000 kg zouden simuleren, zou dit model resulteren in een zwemsnelheid van 1,50 m/s of 5,4 km/uur. Dit model is zeer effectief gebleken, niet alleen bij het berekenen van de zwemsnelheid van de Megalodon, maar ook bij het opstellen van een model voor meerdere haaiensoorten (Fig. 4).
Het is echter de uitbarstingssnelheid die haaien gebruiken om hun prooi snel te grijpen. Witte haaien gebruiken dit voor hun “breaching behaviour”, waarbij ze zichzelf uit het water voortbewegen door prooien van onderaf op te jagen om ze bij verrassing te vangen (fig. 1) . Zij kunnen dit doen omdat zij door tegenschaduw goed gecamoufleerd zijn. Dus, omdat Megalodon waarschijnlijk ook tegenschaduwde, en een vergelijkbare ecologie en lichaamsstructuur had als witte haaien, roept dit de vraag op: was Megalodon ook in staat om te breken?
Misschien. De uitbraaksnelheid van de witte haai wordt aangedreven door een fysiologische aanpassing die mesothermie wordt genoemd. Dit is iets waarvan onlangs is gesuggereerd dat Megalodon het ook had (in detail te bespreken in vraag 5). Mesotherme vissen hebben over het algemeen een hogere snelheid dan ectotherme vissen, en dit is gebruikt om te suggereren dat Megalodon een snelheid van 37.15 km/uur kan hebben gehad.
Die ongelooflijk hoge snelheid voor zo’n grote haai kan goed genoeg zijn om een bres te slaan. Bovendien is de reuzenhaai (Cetorhinus maximus), de op één na grootste haai van vandaag, ook waargenomen terwijl hij doorbrak. Misschien was de reusachtige Megalodon inderdaad in staat om uit het water te springen toen hij zijn laatste maaltijd greep. We weten het gewoon nog niet, maar mijn persoonlijke en (enigszins) professionele mening is dat het heel goed mogelijk zou kunnen zijn geweest.
Met zo’n breed menu om uit te kiezen, was Megalodon een apex roofdier, zonder bekende roofdieren van zichzelf. Dit betekent twee belangrijke dingen. Ten eerste had hij waarschijnlijk zware concurrentie voor zijn prooi, iets dat uiteindelijk kan hebben bijgedragen tot zijn uitsterven. Ten tweede zou Megalodon, als top-predator, van het grootste belang zijn geweest voor het ecosysteem, omdat hij de prooipopulaties in toom zou hebben gehouden en zo een gezond ecosysteem in stand zou hebben gehouden.
Dit is de reden waarom de haaien van vandaag zo belangrijk zijn voor het mariene milieu. Het verlies van een roofzuchtige haai veroorzaakt onomkeerbare cascade-effecten die de gemeenschap veranderen die hij achterlaat. Zo zijn onze mariene ecosystemen deels zoals ze zijn omdat Megalodon er niet langer deel van uitmaakt. En de enorme walvissen van de moderne oceanen kunnen daardoor gerust zijn.
- Estrada JA, Rice AN, Natanson LJ & Skomal GB 2006. Use of isotopic analysis of vertebrae in reconstructing ontogenetic feeding ecology in white sharks. Ecology 87, 829-834.
- Pimiento C, Ehret DJ, MacFadden BJ & Hubbell G 2010. Ancient nursery area for the extinct giant shark Megalodon from the Miocene of Panama. PLoS One 5, e10552.
- Pimiento C, González-Barba G, Ehret DJ, Hendy AJ, MacFadden BJ & Jaramillo C 2013. Sharks and rays (Chondrichthyes, Elasmobranchii) from the late Miocene Gatun formation of Panama. J. Paleontol. 87, 755-774.
- Purdy RW 1996. Paleoecologie van fossiele witte haaien. In: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 67-78, Academic Press, San Diego.
- Aguilera OA, García L & Cozzuol MA 2008. Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation. Paläontol. Z. 82, 204-208.
- Fallows C, Gallagher A. & Hammerschlag N 2013. White sharks (Carcharodon carcharias) scavenging on whales and its potential role in further shaping the ecology of an apex predator. PLoS One, 8, e60797.
- Dudley SFJ, Anderson-Reade MD, Thompson GS & McMullen PB 2000. Concurrent scavenging of a whale carcass by great white sharks, Carcharodon carcharias, and tiger sharks, Galeocerdo cuvier. Fish. Bull. 98, 646-649.
- Kallal RJ, Godfrey SJ & Ortner DJ 2010. Bone reactions on a Pliocene cetacean rib indicate short-term survival of predation event. Int. J. Osteoarchaeol. 22, 253-260.
- Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M & Bianucci G 2017. Had de uitgestorven reuzenhaai Carcharocles megalodon het gemunt op kleine prooien? Bijtsporen op zeezoogdierresten uit het Laat-Mioceen van Peru. Palaeogeogr. Palaeoclimatol. Palaeoecol. 469, 84-91.
- Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, Ferrara TL, Cunningham E, Dean MN & Summers AP 2008. Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite? J. Zool. 276, 336-342.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2015. Is de schaling van zwemsnelheid bij haaien gedreven door metabolisme? Biol. Lett. 11, 20150781.
- Gottfried MD, Compagno LJV & Bowman SC 1996. Size and skeletal anatomy of the giant “megatooth” shark Carcharodon megalodon. In: Grote Witte Haaien: de biologie van Carcharodon carcharias (eds. Klimley AP & Ainley DG), 55-66, Academic Press, San Diego.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2016. Correction to ‘Is the scaling of swim speed in sharks driven by metabolism?’ Biol. Lett. 12, 20160775.
- Watanabe YY, Payne NL, Semmens JM, Fox A & Huveneers C 2019. Zwemstrategieën en energetica van endotherme witte haaien tijdens het foerageren. J. Exp. Biol. 222, jeb185603.
- Cooper JA, Pimiento C, Ferrón HG & Benton MJ (in press). Lichaamsafmetingen van de uitgestorven reuzenhaai Otodus megalodon: Een 2D reconstructie. Sci. Rep.
- Watanabe YY, Goldman KJ, Caselle JE, Chapman DD & Papastamatiou YP 2015. Comparative analyses of animal-tracking data reveal ecological significance of endothermy in fishes. Proc. Nat. Acad. Sci. 112, 6104-6109.
- Ferrón HG 2017. Regionale endothermie als trigger voor gigantisme bij sommige uitgestorven macropredatoire haaien. PLoS One, 12, e0185185.
- Johnston EM, Halsey LG, Payne NL, Kock AA, Iosilevskii G, Whelan B & Houghton JD 2018. Latente kracht van reuzenhaaien onthuld door uitzonderlijke breaching events. Biol. Lett. 14, 20180537.
- Myers RA, Baum JK, Shepherd TD, Powers SP & Peterson CH 2007. Cascading effects of the loss of apex predatory sharks from a coastal ocean. Science, 315, 1846-1850.
Geredigeerd door Rhys Charles
Geef een antwoord