Frontiers in Psychology
On september 28, 2021 by adminHet corpus callosum (CC) is de primaire commissuur die de twee hersenhelften met elkaar verbindt. Hoewel de term commissurotomie niet onjuist is om de zogenaamde “gespleten hersenen” te beschrijven, en strikt correct is voor de oorspronkelijke menselijke chirurgische ingrepen waarbij alle commissuren werden doorgesneden (Gazzaniga et al., 1967), bij latere operaties werd alleen de CC doorgesneden, waardoor de term “callosotomie” in gebruik werd genomen, die in die gevallen de meer passende term blijft (bijv. Franz et al., 1996, 2000).
Uit evolutionair oogpunt is de CC een recente ontwikkeling die alleen bij placentazoogdieren is waargenomen (Katz et al., 1983; Mihrshahi, 2006). De functionaliteit van de CC zou, althans gedeeltelijk, zijn voorafgegaan door de voorste commissuur (AC), zoals waargenomen bij niet-placentale zoogdieren zoals buideldieren (Heath and Jones, 1971; Ashwell et al., 1996). Hoewel sommige overgebleven paden van de AC nog steeds aanwezig zijn in de menselijke hersenen, blijven de connectiviteit en structuur ervan onderbestudeerd in de huidige literatuur en de schaarse bevindingen zijn nogal inconsistent (Lasco et al., 2002; Patel et al., 2010; Choi et al., 2011). Bijvoorbeeld, de bevindingen op basis van magnetische resonantie beeldvorming (MRI) zijn gemengd over de vraag of de AC van vergelijkbare grootte is bij vrouwen en mannen (Lasco et al., 2002; Patel et al., 2010). In een relatief recente studie waarbij gebruik werd gemaakt van de in-vivo techniek van diffusie tensor imaging (DTI) die gebruik maakt van de diffusie van watermoleculen in axonen, onderzochten Patel et al. (2010) acht proefpersonen en isoleerden met succes vezels van het AC bij vijf proefpersonen. Een van de bevindingen was dat bij één proefpersoon een groot deel van de vezels in de achterste ledematen van het AC bilateraal naar de pariëtale gebieden gingen, waar de meeste eindigden, met een kleinere bundel die eindigde in de precentrale gyrus, en nog een andere bundel die de temporale kwab binnenging. Dergelijke individuele verschillen werden gevonden bij alle deelnemers, maar een meerderheid van de proefpersonen toonde bilaterale projecties naar de orbitofrontale, pariëtale en temporale cortex, waar de CC niet duidelijk naar projecteert. Samen impliceren de bevindingen dat de AC zowel primaire als rudimentaire projecties heeft naar neocorticale gebieden waar de CC niet op projecteert, zoals anderen ook hebben gesuggereerd (Di Virgilio et al., 1999; Patel et al., 2010).
In onze visie is de AAA sterk afhankelijk van neocorticale netwerken en connectiviteit met subcorticale structuren zoals de basale ganglia en thalamus (onder andere niet specifiek besproken in ons voorstel) (Franz, 2012). Evolutionaire ontwikkelingen van het AC zouden dus ook een rol kunnen spelen in het AAA model. We hebben echter weinig literatuur gevonden die specifiek ingaat op dit mogelijke verband.
De rol van het AC is bestudeerd in taken die waarschijnlijk vormen van aandacht vereisen, zoals die waarbij visuele aandacht betrokken is. Het is bijvoorbeeld bekend dat acallosale populaties zijn bestudeerd in de context van taken waarbij visuele integratie een rol speelt (Gazzaniga et al., 1962, 1965; Sperry, 1968; Levy et al., 1972). Meer recentelijk heeft Corballis (1995) getracht visuele integratie en aandacht aan de orde te stellen in een poging om eerdere discrepanties in acallosaal onderzoek te verklaren. Corballis stelde een dichotomisch visueel aandachtssysteem voor; een deel (dat betrekking heeft op de vorm) is automatisch en gelokaliseerd in elke hemisfeer, en het andere deel (dat betrekking heeft op beweging en plaats) is vrijwillig en subcorticaal. Het AAA model kan op deze observatie voortbouwen en de in de literatuur waargenomen discrepanties aanpakken. Gebaseerd op ons voorgestelde model, zijn er op meerdere niveaus van aandacht andere routes dan de CC die informatieoverdracht tussen hersenhelften zouden kunnen bemiddelen. Een mogelijkheid is het circuit van de basale ganglia, waarvan wij denken dat het mogelijk cruciaal is voor het delen van aandacht tussen hersenhelften (Franz, 2012). Door het doorsnijden van de CC wordt de directe overdracht van aandacht via corticale paden grotendeels opgeheven (tenzij, natuurlijk, de AC een backup rol speelt). Toch zou de op aandacht gebaseerde integratie van visuele informatie nog steeds gemedieerd kunnen worden via subcorticale paden en/of de AC. Op deze manier zou het tweeledige aandachtssysteem dat door Corballis (1995) is waargenomen, aanwezig kunnen zijn door de aandachtsgebaseerde bemiddeling van visuele integratie via het AC (en/of de basale ganglia en betrokken circuits: Franz, 2012).
Studies waarbij visie en aandacht betrokken zijn, hoeven niet geheel los te worden gezien van het systeem dat motorische actie reguleert. Op basis van onze eigen bevindingen (Franz, 2004; Franz en Packman, 2004) zijn verschillende vormen van aandacht (visueel en intern-niet-visueel) nauw verbonden met motorische actie, en het spreekt voor zich dat een mogelijke sleutel tot het begrijpen van een potentiële aandachtsrol van het AC ligt in de link tussen actie, visie, en aandacht. In een eerdere studie liet Franz (2004) normale deelnemers bimanueel cirkels tekenen onder verschillende manipulaties van visuele feedback, of interne aandacht voor elke hand. De grootte van de cirkels getrokken door een hand die een vorm van interne aandacht kreeg, werd groter op een manier die vergelijkbaar is met effecten die optraden bij de aanwezigheid van visuele feedback van die hand (ten opzichte van geen aandacht/visuele feedback). Ons AAA model impliceert dat aandacht een sleutelcomponent is in de regulatie van motorische output (alsook in de selectie van welke motorische actie gepland is), en voor zover het huidige bewijsmateriaal reikt, kunnen we niet uitsluiten dat het AC een rol speelt in die functie. Een directe test van de rol van het AC in een cognitieve taak is echter niet triviaal.
Testen of het AC een rol speelt in cognitieve functie is bijna onmogelijk in neuropsychologisch onderzoek gezien het feit dat deelnemers met gelokaliseerde AC niet gemakkelijk te vinden zijn (en wij kennen er geen). Het zou daarom veel intuïtiever kunnen zijn om neurologische verschillen in het normale AC te beoordelen en neurologische gegevens te correleren met gedragsgegevens. De gepresenteerde tractografie methode (het ophelderen van het AC) kan relatief gemakkelijk worden toegepast op grote steekproeven van de normale bevolking, en idealiter kan het worden gecombineerd met prestaties (gebaseerd op taken uitgevoerd tijdens afzonderlijke testsessies van de DTI collectie) van diezelfde individuen met behulp van doel-ontworpen cognitieve taken. Hier tonen we aan dat DTI kan worden uitgevoerd en bruikbare resultaten kan opleveren bij een kleine steekproef van mensen uit de neurologisch-normale populatie. Het algemene voorstel zou zijn om soortgelijke methoden uit te voeren op een grotere steekproef, en (zoals hierboven gesuggereerd), correleren bevindingen (van een proxy maatregel van de dichtheid van de AC, bijvoorbeeld) met variabelen op een cognitieve taak.
In figuur 1 tonen we resultaten van DTI tracking op gegevens van 10 vrouwelijke deelnemers verkregen uit een openbare DTI dataset aan de Johns Hopkins University (zie Acknowledgments voor de volledige bron). De gemiddelde leeftijd was 22 jaar (SD = 2,5). Beelden werden verwerkt met behulp van FSL Diffusion Toolbox (Smith et al., 2004; Woolrich et al., 2009; Jenkinson et al., 2012) en procedures in dienst die in eerdere studies gebruikt op andere vezels traktaten (FDT; Behrens et al., 2003a,b, 2007; Johansen-Berg et al., 2004). Kort gezegd, beelden werden eerst samengevoegd tot 4d volumes voor elke deelnemer en schedels gestript met behulp van FSL’s Brain Extraction Tool (BET; Smith, 2002; Jenkinson et al., 2005). Gewonnen beelden werden gecorrigeerd voor kleine hoofd beweging en wervelstromen gegenereerd door veranderingen in de richting van het veld. Diffusie tensoren werden gemonteerd op beelden die primaire diffusie richtingen en niveaus van FA weer te geven op elke voxel te worden gebruikt voor kleurkaarten tijdens het maskeren genereren. Bayesiaanse schatting van diffusie parameters verkregen met behulp van bemonstering technieken met kruising vezels (BEDPOSTX; “X” impliceert kruising vezels) werd uitgevoerd om een verdeling van diffusie grootte en richting rekening houdend met twee kruising vezels om later tractografie uit te voeren over diffusie voxels.
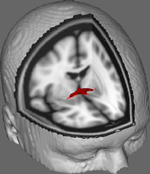
Figuur 1 te genereren. Figuur 1. Gemiddelde trackingresultaten van 10 controledeelnemers, gelegd op een standaard T1-gewogen MRI-beeld met 2 mm isotrope voxels voor anatomische referentie.
Gemiddelde trackingresultaten van alle 10 controledeelnemers werden vervolgens gelegd op een standaard T1-gewogen MRI-beeld met 2 mm isotrope voxels voor anatomische referentie. De groep gemiddelde FA ratio was 0.3146 (SD = 0.0472). Deze tractografie resultaten onthullen, belangrijk, dat de structuur van de AC kan worden opgehelderd, en heeft daarom potentieel voor verdere studies correleren prestaties scores op gedragstaken met FA-waarden met behulp van een grotere steekproefgrootte.
In samenvatting, op dit moment bestaat er een schaarse literatuur, en van de beschikbare literatuur geen specifiek verband tussen de AC en processen met betrekking tot het AAA-model, dat is de focus van onze eerdere doelartikel (Franz, 2012). Met behulp van een procedure vergelijkbaar met die hierin voorgesteld, is het mogelijk om grotere steekproeven van de normale bevolking te testen, en de FA-waarden van de DTI resultaten (bijvoorbeeld) te correleren met een specifieke afhankelijke variabele verkregen uit een doel-ontwikkelde taak van aandacht voor actie (procedures waarvan de ontwikkeling in ons laboratorium gaande is).
Conflict of Interest Statement
De auteurs verklaren dat het onderzoek werd uitgevoerd in de afwezigheid van enige commerciële of financiële relaties die zouden kunnen worden opgevat als een potentieel belangenconflict.
Acknowledgments
Wij erkennen dankbaar het Human Brain Project en National Research Resource Center (USA) en John Hopkins University voor het toestaan van openbaar gebruik van hun DTI-gegevens. We zijn ook dankbaar voor gulle subsidie steun van de Marsden Fonds van de Royal Society of New Zealand (aan Elizabeth A. Franz).
Corballis, M. (1995). Visuele integratie in het gespleten brein. Neuropsychologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). The allocation of attention to learning of goal-directed actions: a cognitive neuroscience framework focusing on the basal ganglia. Front. Psychol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., and Jones, E. (1971). Interhemisferische paden bij afwezigheid van een corpus callosum. Een experimentele studie van commissurale connexies in het buideldier phalanger. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
Geef een antwoord