Núcleo olivariano superior
On Novembro 12, 2021 by admin| Cérebro: Complexo olivariano superior | ||
|---|---|---|
 |
||
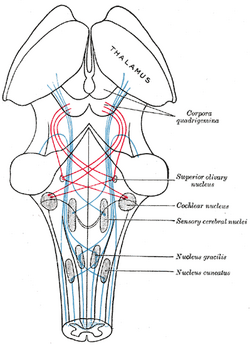
| Esquema mostrando o curso das fibras do lemnisco; lemnisco medial em azul, lateral em vermelho. (Núcleo olivariano superior é rotulado no centro direito.) | ||
| ] | ||
| Latina | núcleo olivaris superior | |
| Cinza | assunto #187 787> | |
| Parte de | ||
| Componentes | ||
| Artery | ||
| Vein | ||
| BrainInfo/UW | {{{BrainInfoType}}}-565 | |
| MeSH | ||
O complexo olivariano superior (ou SOC ou oliva superior) é uma pequena massa de substância cinza situada na superfície dorsal da parte lateral do corpo trapezoidal. Pequeno em humanos, mas bem desenvolvido em certos animais, apresenta estrutura semelhante ao núcleo olivariano inferior, e está situado imediatamente acima dele. Algumas das fibras do corpo trapezoidal terminam por arborizar ao redor das células deste núcleo, enquanto outras surgem destas células.
Fisiologia
O núcleo olivariano superior desempenha uma série de papéis na audição. O olivo superior medial (MSO) é um núcleo especializado que se acredita medir a diferença temporal da chegada dos sons entre os ouvidos (a diferença temporal interaural ou ITD). O ITD é uma pista importante para determinar o azimute dos sons de baixa frequência, ou seja, localizá-los no plano azimutal – o seu grau à esquerda ou à direita.
A oliveira superior lateral (LSO) acredita-se estar envolvida na medição da diferença de nível de sons entre os ouvidos (a diferença de nível interaural ou ILD). A DPI é uma segunda pista principal na determinação do azimute dos sons de alta freqüência.
Relação com o sistema auditivo
O complexo olivar superior está geralmente localizado nas pons, mas no humano estende-se da medula rostral até as mid-pons e recebe projeções predominantemente do núcleo coclear anteroventral através das estrias acústicas ventrais, embora o núcleo póstero-ventricular também se projeta ali através das estrias acústicas intermediárias. É neste local que ocorrem as primeiras grandes interacções binaurais.
Núcleos primários
O complexo olivariano superior é dividido em três núcleos primários, o MSO, LSO, e o Núcleo Medial do corpo trapezoidal, e vários núcleos periolivares menores. Estes três núcleos são os mais estudados e, portanto, melhor compreendidos. Tipicamente, eles são considerados como formando o caminho de localização do azimute ascendente.
Olive superior medial (MSO)
Pensa-se que a oliveira superior medial ajuda a localizar o azimute de um som, ou seja, o ângulo à esquerda ou à direita onde a fonte sonora está localizada. O primeiro instinto pode ser pensar que este núcleo inclui informação vertical, mas não é o caso; a informação processada no MSO é restrita ao plano horizontal. As células fusiformes não se projetam a nada no nível dos pons, e só entram em jogo no colículo inferior. Apenas dados horizontais estão presentes, mas vêm de duas fontes auditivas diferentes, o que ajuda na localização do som no eixo do azimute. A forma como a azeitona superior o faz é medindo as diferenças no tempo entre dois sinais auditivos que gravam o mesmo estímulo. Percorrer a cabeça leva cerca de 700 μs, e supõe-se que a azeitona superior medial é capaz de detectar isto. De fato, observa-se que as pessoas podem detectar diferenças interauriculares de até 10 microssegundos. O núcleo está tonotopicamente organizado, mas a projecção do campo receptivo azimutal é “muito provavelmente um mapa complexo, não linear”.
As projecções da oliveira superior medial terminam densamente no núcleo central ipsilateral do colículo inferior. A maioria destes axônios são considerados de “forma redonda” ou tipo R. Estes axônios R são na sua maioria glutamátricos e contêm vesículas sinápticas redondas e formam junções sinápticas assimétricas.
- Este é o maior dos núcleos e em humanos contém aproximadamente 15.500 neurônios .
- Cada MSO recebe entradas bilaterais de baixa freqüência dos AVCNs direito e esquerdo.
- A saída é para o lêmniscus lateral ipsilateral e finalmente para o colículo inferior.
- O MSO responde melhor aos estímulos binaurais.
- A função principal do MSO envolve a detecção de diferenças no tempo de chegada dos sons aos dois ouvidos, que faz parte do processo de localização (ITD).
Azeitona superior lateral (LSO)
Esta azeitona tem funções semelhantes à azeitona superior medial, mas emprega intensidade para localizar a fonte sonora. Esta é a parte do tronco cerebral que rotula o som mais alto do ouvido esquerdo como estando no lado esquerdo. A azeitona lateral recebe a entrada de ambos os núcleos cocleares, das células arbustivas ipsilaterais do AVCN esférico e das células arbustivas globulares contralaterais do AVCN através da MNTB ipsilateral. As células principais MNTB são glicinérgicas, fornecendo uma fonte de inibição para o LSO, e formando a base da sensibilidade ILD. As projeções de ambos os núcleos cocleares são primariamente de alta freqüência, e essas freqüências são posteriormente representadas pela maioria dos neurônios LSO (>2/3 acima de 2-3kHz em gato). Entradas adicionais derivam do LNTB ipsilateral (glicinérgico, veja abaixo), que fornece informações inibitórias a partir do núcleo coclear ipsilateral. Outro input possivelmente inibitório deriva das células ipsilaterais não esféricas do AVCN. Estas células ou são globulares ou multipolares (estelato). Qualquer um destes dois inputs poderia fornecer a base para a inibição ipsilateral vista em mapas de resposta flanqueando a excitação primária, afiando o ajuste de freqüência da unidade.
O LSO projeta bilateralmente para o núcleo central do colículo inferior (ICC). As projeções Ipsilateral são principalmente inibitórias (glicinérgicas), e contralaterais excitatórias. Outros objetivos de projeção incluem os núcleos Dorsal e Ventral do Lemniscus Lateral (DNLL & VNLL). As projeções GABAergicas do DNLL formam a fonte primária do GABA no tronco cerebral auditivo, e projetam bilateralmente para o ICC e para o DNLL contralateral. Essas conexões excitatórias e inibitórias convergentes podem atuar para diminuir o nível de dependência da sensibilidade ao DPI no ICC comparado ao LSO.
Projeções adicionais formam o feixe Olivococlear Lateral (LOC), que inerva as células capilares internas cocleares. Pensa-se que estas projecções têm uma constante de tempo longa, e actuam para normalizar o nível sonoro detectado por cada ouvido de forma a ajudar na localização do som. Existem diferenças consideráveis entre as espécies: Os neurônios de projeção LOC são distribuídos dentro do LSO em roedores, e envolvem o LSO em predadores (ou seja, gato).
Núcleo Medial do Corpo Trapezoidal (MNTB)
- O MNTB é a menor parte do SOC; a presença do MNTB no tronco cerebral humano está em dúvida.
- Cada neurónio recebe apenas uma entrada de alta frequência através de um Calyx de segurado do AVCN contralateral, e os seus projectos de saída para o LSO ipsilateral.
- Existem dois tipos de resposta encontrados: um ‘tipo picador’ semelhante às células do fuso no AVCN e um tipo primário semelhante às células do fuso no AVCN.
Núcleos Periolivares
O SOC é composto por entre seis e nove núcleos periolivares, dependendo do pesquisador citado, tipicamente nomeados com base na sua localização em relação aos núcleos primários. Estes núcleos envolvem cada um dos núcleos primários, e contribuem tanto para o sistema auditivo ascendente como descendente. Estes núcleos também formam a fonte do feixe olivococlear, que inerva a cóclea. Na cobaia, as projeções ascendentes para os colículos inferiores são primariamente ipsilateral (>80%), sendo a maior fonte única proveniente do SPON. Também, os núcleos ventrais (RPO, VMPO, AVPO, & VNTB) são quase inteiramente ipsilaterais, enquanto os núcleos restantes se projetam bilateralmente.
| Nome | Cat | Guinea Pig | Rato | Mouse | |
|---|---|---|---|---|---|
| LSO | X | X | X | X | X |
| MSO | X | X | X | X | X |
| MNTB | X | X | X | X | X |
| LNTB | X | X | “LVPO” | X | |
| ALPO | X | X | > | ||
| PVPO | X | X | |||
| PPO | X | X | X | “CPO” | |
| VLPO | X | > | |||
| DPO | X | X | X | X | |
| DLPO | X | X | > | > | |
| VTB | X | X | “MVPO” | X | |
| AVPO | X | > | > | ||
| VMPO | X | X | > | ||
| RPO | X | X | |||
| SPN | “DMPO” | X | X | X |
Núcleo Ventral do Corpo Trapezoidal (VNTB)
- O VNTB é um pequeno núcleo localizado lateralmente ao MNTB, e ventral para o MSO.
- Composto de uma população heterogênea de células, este núcleo se projeta para muitos núcleos auditivos, e forma o feixe olivococlear medial (MOC) que inerva as células capilares externas cocleares. Estas células contêm fibras eletromoleculares, e atuam como amplificadores/atenuadores mecânicos dentro da cóclea.
- O núcleo projeta para ambos os CI, sem células projetando bilateralmente.
Núcleo lateral do corpo trapezoidal (LNTB)
- Ventral localizado ao LSO
- Células arbustivas esféricas AVCN projetam colaterais bilateralmente, e células arbustivas globulares projetam colaterais ipsilateralmente aos neurônios LNTB.
- As células são imunoreativas para glicina, e são rotuladas retrogradadamente após a injeção de glicina tritiada no LSO
- O núcleo projeta tanto para a CI, com poucas células projetando bilateralmente, quanto para o LSO ipsilateral.
- Células multipolares grandes projetam o núcleo coclear, mas não a CI, tanto na gata quanto na cobaia.
- Entradas são frequentemente através de bulbos terminais de retenção, produzindo transdução de sinal muito rápido.
Núcleo Periolivar Superior (SPON) (Núcleo Periolivar Dorsomedial (DMPO))
- Locado directamente dorsal ao MNTB
- Em rato, SPON é um núcleo GABAérgico homogéneo. Estes neurônios tonotopicamente organizados recebem entradas excitatórias de polvo e células multipolares no núcleo coclear ventral contralateral, uma entrada glicinérgica (inibitória) do MNTB ipsilateral, uma entrada GABAergic (inibitória) desconhecida, e projetam-se para o ICC ipsilateral. A maioria dos neurônios reage apenas no desvio de um estímulo, pode bloquear fase a estímulos AM até 200 Hz, e pode formar a base para a seletividade da duração do ICC. Notavelmente, os neurônios SPON não recebem entradas descendentes do CI, e ele não se projeta para a cóclea ou núcleo coclear, como muitos núcleos periolivares fazem.
- Em cobaia, células multipolares redondas a ovais projetam para ambas CI, com muitas células projetando bilateralmente. As células mais alongadas que se projetam para o núcleo coclear não se projetam para o ICC. Parece haver para as populações de células, uma que projeta ipsilateralmente, e outra que projeta bilateralmente.
- A maioria das informações tinha vindo de SPON roedor, devido ao tamanho proeminente do núcleo nestas espécies, com muito poucos estudos foram feitos em DMPO gato, nenhum dos quais foi extenso.
Núcleo Periolivar Dorsal (DPO)
- Localizado dorsal e medial ao LSO
- Contém ambas as unidades EE (excitado por ambas as orelhas) e E0 (excitado apenas pela orelha contralateral).
- Neurões são tonotopicamente organizados, e de alta frequência.
- Pode pertencer a um único núcleo juntamente com o DLPO
- O núcleo projecta-se para ambos os CI, sem células a projectarem-se bilateralmente.
Núcleo Periolivar Dorsolateral (DLPO)
- Localizado dorsal e lateral ao LSO
- Contém ambas as unidades EE (excitado por ambas as orelhas) e E0 (excitado apenas pela orelha contralateral).
- Neurões são organizados tonotopicamente, e baixa frequência.
- Pode pertencer a um único núcleo juntamente com o DPO
- O núcleo projecta-se para ambos os CI, com poucas células a projectarem-se bilateralmente.
Núcleo Ventrolateral Periolivar (VLPO)
- Localizado ventral para e dentro da colina ventral do LSO
- Contém ambas as unidades EI (excitado pelo contralateral e inibido pela orelha ipsilateral) e E0 (excitado apenas pela orelha contralateral).
- Neurões são tonotopicamente organizados, e de alta frequência.
Núcleo Periolivar Anterolateral (ALPO)
- O núcleo projecta-se para ambos os CI, sem células a projectarem-se bilateralmente.
- As células multipolares grandes projectam-se para o núcleo coclear, mas não para o CI, tanto no gato como no porquinho-da-índia.
Núcleo Periolivar Ventromedial (VMPO)
- Localizado entre o MSO e o MNTB.
- Enviar projecções para o ICC bilateralmente.
- O núcleo projecta para ambos os CI, sem células a projectarem-se bilateralmente.
Núcleo Periolivar Rostral (RPO) (Núcleo Periolivar Anterior (APO))
- Localizado entre o pólo rostral do MSO e o VNLL
Núcleo Periolivariano Caudal (CPO) (Núcleo Periolivariano Posterior (PPO))
- Localizado entre o pólo caudal do MSO e o núcleo facial (7N)
Núcleo Periolivar Posteroventral (PVPO)
- O núcleo projecta-se para ambos os CI, sem células a projectarem-se bilateralmente.
Fisiopatologia
Uma autópsia de uma mulher de 21 anos com autismo, epilepsia e retardo mental encontrou uma ausência quase completa da azeitona superior.
Ver também
- Olivary body
- Kulesza RJ, Cytoarchitecture of the human superior olivary complex: Medial and lateral superior olive. Hearing Research 225(2007) 80-90
- 2.0 2.1 Oliver DL, et al. Projecções axonais da azeitona superior lateral e medial até ao colículo inferior do gato: um estudo utilizando a autoradiografia microscópica electrónica. J Comp Neurol. 1995 Set 11;360(1):17-32 Cite o erro: Inválido
<ref>tag; nome “R4” definido várias vezes com conteúdo diferente - Cajal, S. R. Y. e L. Azoulay (1909). Histologie du système nerveux de l’homme et des vertébrés. Paris, Maloine.
- 4.0 4.1 Kandel, et al Principles of Neuroscience. Quarta ed. pp 591-624. Copyright 2000, por McGraw-Hill Co. Erro de citação: Inválido
<ref>tag; nome “R2” definido várias vezes com conteúdo diferente - Kulesza, R. J., Jr. (2007). “Cytoarchitecture of the human superior olivary complex: medial and lateral superior olive”. Res 225(1-2): 80-90.
- Tsuchitani, C. e J. C. Boudreau (1967). “Codificação da frequência e intensidade do estímulo pelas células do segmento S superior da azeitona do gato”. J Acoust Soc Am 42(4): 794-805.
- 8.0 8.1 8.2 Citar erro: Inválido
<ref>tag; nome “R10” definido várias vezes com conteúdo diferente - Wu, S. H. e J. B. Kelly (1994). “Evidência fisiológica para inibição ipsilateral na azeitona superior lateral: respostas sinápticas na fatia cerebral do rato”. Res 73(1) auditivo: 57-64.
- Brownell, W. E., P. B. Manis, et al. (1979). “Ipsilateral inhibitory responses in the cat lateral superior olive.” Resina Cerebral 177(1): 189-93.
- Darrow, K. N., S. F. Maison, et al. (2006). “Cochlear efferent feedback equilibra a sensibilidade interaural”. Nat Neurosci 9(12): 1474-6.
- Warr, W. B. e J. J. J. Guinan, Jr. (1979). “Inervação eferente do órgão de corti: dois sistemas separados”. Resina Cerebral 173(1): 152-5.
- 13.0 13.1 Schofield, B. R. e N. B. Cant (1991). “Organização do complexo olivariano superior no porquinho-da-índia. I. Cytoarchitecture, cytochrome oxidase histochemistry, and dendritic morphology”. J Comp Neurol 314(4): 645-70. Erro de citação: Inválido
<ref>tag; nome “R7” definido várias vezes com conteúdo diferente - 14,0 14,1 14,2 14,3 14,4 14,5 14,6 14,7 14,8 Illing, R. B., K. S. Kraus, et al. (2000). “Plasticidade do complexo olivariano superior”. Microsc Res Tech 51(4): 364-81. Erro de citação: Inválido
<ref>tag; nome “R5” definido várias vezes com conteúdo diferente Erro de citação: Inválido<ref>tag; nome “R5” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R5” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R5” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R5” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R5” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R5” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R5” definido várias vezes com conteúdo diferente - Warr, W. B. e J. E. Beck (1996). “Projecções múltiplas do núcleo ventral do corpo do trapézio no rato”. Res 93(1-2): 83-101.
- 16,00 16,01 16,02 16,03 16,04 16,05 16,06 16,07 16,08 16,09 16,10 Schofield, B. R. e N. B. Cant (1992). “Organização do complexo olivariano superior na cobaia: II. Padrões de projeção dos núcleos periolivares para o colículo inferior”. J Comp Neurol 317(4): 438-55. Cite erro: Inválido
<ref>tag; nome “R6” definido várias vezes com conteúdo diferente Erro de citação: Inválido<ref>tag; nome “R6” definido várias vezes com conteúdo diferente Erro de citação: Inválido<ref>tag; nome “R6” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R6” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R6” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R6” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R6” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R6” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R6” definido várias vezes com conteúdo diferente Erro de dita: Inválido<ref>tag; nome “R6” definido várias vezes com conteúdo diferente - Smith, P. H., P. X. Joris, et al. (1993). “Projections of physiologically characterized spherical bushy cell axons from the cochlear nucleus of the cat: evidence for delay lines to the medial superior olive”. J Comp Neurol 331(2): 245-60.
- Wenthold, R. J., D. Huie, et al. (1987). “Glycine immunoreactivity located in the cochlear nucleus and superior olivary complex.” Neurociência 22(3): 897-912.
- 19.0 19.1 Adams, J. C. (1983). “Cytology of periolivary cells and the organization of their projections in the cat.” (Citologia das células periolivares e a organização de suas projeções no gato). J Comp Neurol 215(3): 275-89. Cite error: Inválido
<ref>tag; nome “R8” definido várias vezes com conteúdo diferente - Friauf, E. e J. Ostwald (1988). “Projeções divergentes dos neurônios do núcleo coclear ventral do rato caracterizados fisiologicamente como mostrado por injeção intra-axonal de peroxidase de rábano silvestre”. Exp Brain Res 73(2): 263-84.
- Kulesza, R. J., Jr. e A. S. Berrebi (2000). “Núcleo superior paraolivary do rato é um núcleo GABAergic.” J Assoc Res Otolaryngol 1(4): 255-69.
- Kulesza, R. J., Jr., G. A. Spirou, et al. (2003). “Propriedades fisiológicas de resposta dos neurônios no núcleo paraolivar superior do rato”. J Neurofisiol 89(4): 2299-312.
- WHITE JS, WARR WB. A dupla origem do feixe olivococlear na ratazana albina. J. Comp. Neurol. 219:203–214, 1983.
- Guinan, J. J. J., Jr., S. S. Guinan, et al. (1972). “Unidades auditivas únicas no complexo olivariano superior. I. Respostas a sons e classificações baseadas em propriedades fisiológicas”. Int J Neurosci 4: 101-20.
- Tsuchitani, C. (1977). “Organização funcional de grupos de células laterais do complexo olivariano superior do gato”. J Neurofisiol 40(2): 296-318.
- Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996). Origem embriológica para o autismo: anomalias de desenvolvimento dos núcleos motores do nervo craniano. J Comp Neurol 370 (2): 247-61.
- MedEd at Loyola Neuro/frames/nlBSs/nl27fr.htm
- Image at med.utah.edu (pick 9b. Pons)
Este artigo foi originalmente baseado em uma entrada de uma edição de domínio público de Gray’s Anatomy. Como tal, algumas das informações contidas aqui podem estar desatualizadas. Por favor, edite o artigo se este for o caso, e sinta-se à vontade para remover este aviso quando ele não for mais relevante.
|
v-d-e
Cérebro: rhombencephalon (hindbrain) |
||||||
|---|---|---|---|---|---|---|
| Metencephalon/ pons |
|
Dorsal/
(tegmentum) |
Ventral/
(base) |
|||
| Myelencephalon/ medulla |
|
Dorsal
|
Ventral
|
Raphe/reticular
|
||
|
v-d-e
Sistema sensorial: Sistema auditivo e Vestibular (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ouvido externo |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Lóbulo da orelha) – Canal auditivo – Músculos auriculares Eardrum (Umbo, Pars flaccida) |
|||||||||||
| Orelha média |
|
|||||||||||
| Inner ear/ (membranous labyrinth, labirinto de carbono) |
|
|||||||||||
| {| class=”navbox colapsable nowraplinks” style=”margin:auto; “ | ||||||||||||
|
—
|
||||||||||||
|
||||||||||||
|}
Esta página usa o conteúdo Creative Commons licenciado da Wikipedia (ver autores).
Deixe uma resposta