N-結合型糖鎖
On 12月 18, 2021 by admin
N-結合型糖鎖の生合成は3つの主要なステップを経て行われる。
- ドリコル型糖鎖の合成
- タンパク質への糖鎖の転移
- 糖鎖の処理
前糖の合成、転移および初期トリミングは小胞体で行われ、前糖は小胞体で合成され、小胞体でトリミングされる。
このように糖タンパク質の合成は、異なる細胞区画で空間的に分離されている。 そのため、合成される糖鎖の種類は、これらの細胞区画内に存在する異なる酵素に対するそのアクセス性に依存する。
しかし、多様性にもかかわらず、すべてのN-グリカンは共通のコア糖鎖構造を持つ共通の経路を通じて合成される。コア糖鎖構造は、本質的に2個のN-アセチルグルコサミンと3個のマンノース残基から構成されている。 このコア糖鎖はその後さらに精緻化され修飾され、多様なN-グリカン構造を生み出す。
前駆体オリゴ糖の合成 編集
N-結合型糖鎖形成のプロセスは、ドリコール結合型GlcNAc糖の生成から開始される。 ドリコールはイソプレン単位の繰り返しからなる脂質分子である。 この分子はERの膜に付着していることが分かっている。 糖分子はピロリン酸結合でドリコールに結合している(1つのリン酸はもともとドリコールに結合しており、2つ目のリン酸はヌクレオチド糖から由来している)。 この前駆体オリゴ糖の組み立ては2つの段階で行われる。 第I相と第II相である。
タンパク質に転移される前駆体分子は、2個のGlcNAc、9個のマンノース、3個のグルコースから構成され、細胞質側ではフェーズIが、内腔側ではフェーズIIが行われる。

|
|
| Phase II | |
はマンノース残基供与体(生成 : Dol-P + GDP-Man → Dol-P-Man + GDP)、Dol-P-Glucはグルコース残基供与体(生成 : Dol-P + UDP-Glc → Dol-P-Glc + UDP)である。
|
|
糖鎖のタンパク質への転移編集
前駆体オリゴ糖が形成されると、完成した糖鎖はER膜内腔で新生ポリペプチドに移動される。 この反応は、ドリコール-糖鎖分子間のピロリン酸結合の切断から放出されるエネルギーによって駆動される。糖鎖が新生ポリペプチドに転移される前に満たすべき条件が三つある:
- アスパラギンは一次構造中の特定のコンセンサス配列(Asn-X-SerあるいはAsn-X-Thrあるいはまれに Asn-X-Cys )に位置している必要がある。
- アスパラギンはタンパク質の三次元構造の中で適切な位置になければならない(糖は極性分子なので、タンパク質の表面にあるアスパラギンに結合する必要があり、タンパク質内に埋没してはならない)
- N-結合型グリコシル化が開始されるためには、小胞体の内側でアスパラギンが見つかることが必要である。
オリゴ糖転移酵素はコンセンサス配列の認識と、小胞体内腔で翻訳されているポリペプチド受容体に前駆体糖鎖を転移する役割を担う酵素である。 したがって、N-結合型糖鎖形成は共翻訳的なイベントである
糖鎖のプロセシング 編集

N-糖鎖プロセシングは小胞体とゴルジ体において行われる。 前駆体分子の最初のトリミングは小胞体で行われ、その後の処理はゴルジ体で行われる。
完成した糖鎖を新生ポリペプチドに転移する際に、2個のグルコース残基が構造から除去される。 グリコシダーゼと呼ばれる酵素がいくつかの糖残基を除去する。 これらの酵素は水分子を利用してグリコシド結合を切断することができる。 これらの酵素は、糖鎖の非還元末端に位置する単糖残基のみに作用するため、エキソグリコシダーゼと呼ばれる。 この最初のトリミング工程は、ERにおいてタンパク質のフォールディングを監視する品質管理工程として機能していると考えられる。
タンパク質が正しくフォールディングされると、グルコシダーゼIとIIにより2つのグルコース残基が除去される。 最後の3番目のグルコース残基の除去は、糖タンパク質が小胞体からシスゴルジ体へ移行する準備ができたことを知らせるものである。 ERマンノシダーゼはこの最後のグルコースの除去を触媒する。 しかし、タンパク質が正しく折りたたまれないと、グルコース残基が除去されず、その結果、糖タンパク質は小胞体を出ることができない。 シャペロンタンパク質(カルネキシン/カルレティキュリン)は、折り畳まれていないタンパク質や部分的に折り畳まれたタンパク質に結合し、タンパク質の折り畳みを助ける。
次のステップでは、シスゴルジでさらに糖残基の付加と除去を行う。 これらの修飾はそれぞれグリコシルトランスフェラーゼとグリコシダーゼによって触媒される。 シスゴルジでは、一連のマンノシダーゼがα-1,2結合の4つのマンノース残基の一部または全部を除去する。 一方、ゴルジの内側では、糖転移酵素がコア糖鎖構造に糖残基を付加し、高マンノース、ハイブリッド、複合糖鎖という3つの主要なタイプの糖鎖を生み出す。
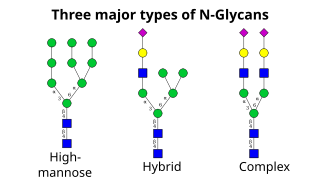
- 高マンノースは、要するに多くのマンノース残基を持つ2つのN-アセチルグルコサミンだけで、しばしばそれがタンパク質に結合する前の前駆体オリゴ糖に見られるのとほぼ同数のマンノース残基が存在する。
- 複合型オリゴ糖は、元の2つ以上のN-アセチルグルコサミンを含む他のタイプの糖のほとんどすべての数を含むことができるので、そのように名付けられた。
- ハイブリッド型オリゴ糖は、マンノース残基を分岐の片側に含み、反対側ではN-アセチルグルコサミンが複合分岐を開始させる。
成長する糖鎖に糖が付加される順序は、酵素の基質特異性と分泌経路を移動する際の基質へのアクセスによって決定される。
ゴルジ体の酵素編集
ゴルジ体の酵素は、様々な種類の糖鎖の合成を決定する上で重要な役割を担っている。 酵素の作用の順番はゴルジ体の積み重ねの中での位置によって反映される。
| 酵素 | ゴルジ体内の位置 | |||||||
|---|---|---|---|---|---|---|---|---|
| マンノシダーゼI | cis->
古細菌と原核生物で編集原核生物と古細菌で同様の糖鎖生合成経路が発見された。 しかし、真核生物と比較して、真正細菌や古細菌の最終的な糖鎖構造は、小胞体で作られる最初の前駆体とあまり変わらないようである。 真核生物では、最初の前駆体オリゴ糖は細胞表面に到達するまでに広範囲に修飾される 。 |
コメントを残す