Gompertz function
On 12月 12, 2021 by adminGompertz curveEdit
Population biologyは特にGompertz関数に関係しています。 この関数は、ある生物の集団の急速な成長を記述するのに特に有用であり、また、環境収容力が決定されると、最終的に水平漸近を説明することができます (プラトー細胞/集団数):
これは次のようにモデル化されます:

ここで。
- tは時間
- N0は細胞の初期量
- NIはプラトー細胞/集団数
- bは腫瘍成長の初期速度
プラトー細胞数のこの関数考慮によって、実際の集団力学を正確に真似ることに有用である。 またこの関数は、一般に集団の成長を詳述する際に最も広く受け入れられているシグモイド関数に忠実である。 さらに、この関数では、対数相を経て急激に数を増やす細菌細胞や癌細胞の集団でよく見られる初期成長率を利用している。 しかし、腫瘍の初期増殖率という関数は、患者さんの小宇宙や、集団生物学でいうところの環境因子が様々であるため、事前に決定することは困難である。 がん患者の場合、年齢、食事、民族性、遺伝的素因、代謝、ライフスタイル、転移の起源などの要因が、腫瘍増殖速度の決定に関与している。 キャリングキャパシティもこれらの要因に基づいて変化すると考えられるので、そのような現象を記述するのは難しい。
Metabolic curveEdit
代謝機能は特に生物内の代謝速度を説明することに関係するものである。 この関数は、腫瘍細胞を監視するために適用することができます。代謝率は動的であり、非常に柔軟であるため、癌の成長を詳述する上でより正確にすることができます。 代謝曲線は、組織が維持され、作られる際に身体が提供するエネルギーを考慮に入れています。 このエネルギーは代謝とみなすことができ、細胞分裂において特定のパターンに従っています。 エネルギー保存は、質量や発生時間の違いに関係なく、このような成長をモデル化するために使用することができます。 すべての分類群に類似の成長パターンがあり、このモデルは、結果として、腫瘍の発生の基礎となる細胞分裂を考慮しています。

- B =生物が安静時に使うエネルギー
- NC =与えられた生物の細胞の数
- BC =個々の細胞の代謝率
- NCBC =現存するものを保つのに必要なエネルギー 組織
- EC= 個々の細胞から新しい組織を作るのに必要なエネルギー
安静時に使われるエネルギーと代謝率の仕事を区別することで、モデルはより正確に成長速度を決定することができます。 安静時のエネルギーは組織を維持するために使われるエネルギーよりも低く、共に既存の組織を維持するために必要なエネルギーを表しています。
腫瘍の成長編集
1960年代にレアードが初めてゴンペルツ曲線を用いて腫瘍の成長データをフィットさせることに成功した。 実際、腫瘍は栄養の供給が制限された狭い空間で成長している細胞集団である。 腫瘍の大きさをX(t)とすると、ゴンペルツ曲線は次のように書くと便利である:
X ( t ) = K exp ( log ( X ( 0 ) K ) exp ( – α t ) ) {displaystyle X(t)=K}exp \left({}frac {X(0)}{K}}right)\exp \left(-┣ㅂ)}

ここで、
- X(0) は観測開始時刻の腫瘍サイズ、
- K は担体容量、すなわち、(a)は(b)を表します。すなわち、利用可能な栄養素で到達可能な最大サイズである。 実際には

独立に X(0)>0.Note that, in absence of therapies etc… .
- α は細胞の増殖能力に関係する定数で、
- log() は自然対数を意味する。
X(t)のダイナミクスはGompertz微分方程式によって支配されることが示される:

すなわち、分解すると次のような形になる。

F(X) は細胞集団の瞬間的な増殖率である。 の減少する性質は、ロジスティック成長率と同様に、細胞数の増加による栄養分の奪い合いによるものである。 しかし根本的な違いがあって、ロジスティックな場合、小さな細胞集団の増殖率は有限である:

ここで、Gompertzの場合、増殖率は無制限である。

SteelやWheldonが気づいたように、細胞集団の増殖速度は結局、細胞分裂時間に束縛される。 したがって、このことはゴンペルツ方程式が小さな腫瘍の成長をモデル化するのに適していないことの証拠かもしれない。 さらに、最近では、免疫系との相互作用を含めて、ゴンペルツや他の非有界F(0)を特徴とする法則は、免疫監視の可能性を排除することが注目されている
Fornalskiらの理論研究は、放物線関数がより適切なごく初期の段階を除いて、がん成長に対するゴンペルツ曲線の生物物理的基礎を示した。
Gompertz growth and logistic growthEdit
The Gompertz differential equation
は一般化ロジスティック微分方程式

(where ν > 0 {displaystyle \nu >0}})

は正の実数)より

.
おまけ。 一般化ロジスティック関数のグラフに変曲点が存在するのは

となり

.
COVID-19感染軌跡のモデリング編集

疫学モデリングにおける一般化ロジスティック関数(リチャード成長曲線)
リチャード成長曲線とも呼ばれる一般化ロジスティック関数は、COVID-19感染軌跡のモデリングに広く使用されています。 感染軌跡とは、国、都市、州などの対象について、累積感染者数の日次時系列データである。 文献上では様々な再パラメータ化が行われており、よく用いられる形式として
f ( t ; θ 1 , θ 2 , θ 3 , ξ ) = θ 1 1 / ξ {displaystyle f(t;\theta _{1},\theta _{2},\theta _{3},\xi )={hank {THEATA _{1}{^{1/entaxi }}}}がある。}

where θ 1 , θ 2 , θ 3 {\displaystyle \theta _{1},\theta _{2},\theta _{3}}}.

は実数、ξ {displaystyle \xi } は実数。

は正の実数である。 曲線f{displaystyle f}の柔軟性

はパラメータ ξ {displaystyle \xi } に起因するものである。
: (i) if ξ = 1 {displaystyle \xi =1} {displaystyle \xi =1}.

ならば曲線はロジスティック関数に還元され、(ii) ξ {displaystyle \xi } ならば、曲線はロジスティック関数に還元される。
が0に収束すると、曲線はGompertz関数に収束する。 疫学モデリングでは、θ 1 {displaystyle \theta _{1}} 。

, θ 2 {displaystyle \theta _{2}}} 。

, and θ 3 {displaystyle \theta _{3}}.

はそれぞれ最終的な流行規模、感染率、ラグフェーズを表す。 右図は、( θ 1 , θ 2 , θ 3 ) {displaystyle (\theta _{1},\theta _{2},\theta _{3})} における感染軌跡の例である。

は、( 10 , 000 , 0.2 , 40 ) {displaystyle (10,000,0.2,40)} で指定されます。

.
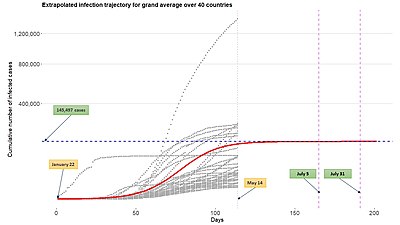
疫学モデリングで一般化ロジスティック関数などの成長関数を用いる利点は、成長関数で複数の対象(国、地域)からの感染経路の記述によりマルチレベルモデルの枠組みに比較的容易に展開できることである。 市、州など)。 上図参照 このようなモデリングの枠組みは、広く非線形混合効果モデルや階層的非線形モデルとも呼ばれる。
Gomp-ex law of growthEdit
以上の考察をもとに、WheldonはGomp-Exモデルという、Gomperts法則を少し修正した腫瘍成長の数学モデルを提案した。 Gomp-Exモデルでは、当初は資源をめぐる競争がないため、細胞集団は指数関数的な法則に従って拡大すると仮定されている。 しかし、臨界サイズ閾値X C {displaystyle X_{C}} が存在する。

such that for X > X C {displaystyle X>X_{C}}.

. 資源をめぐる競争がないという仮定は、ほとんどのシナリオで成立する。 しかし、それは制限要因によって影響される可能性があり、それには副要因変数の作成が必要です。
成長はGompertzの法則に従う:

so that:

ここで、X C {displaystyle X_{C}}の数値的な推定がある。
コメントを残す