細菌の構造と機能
On 10月 10, 2021 by adminStructure and Function of Bacterial Cells (page 5)
(This chapter has 10 pages)
© Kenneth Todar, PhD
Cell Wall
The cellwalls of bacteria deserve specialattention for several reasons:
1.細菌は、細胞壁を持つ。 細胞壁は、前述のように生存に不可欠な構造である。
2.自然界ではどこにも見られないユニークな成分で構成されている。
3.抗生物質による攻撃を受ける最も重要な部位の1つである。 細胞壁は、細胞膜を機械的な損傷や浸透圧による破裂・溶解から守る重要な構造である。原核生物は通常、比較的希薄な環境に生息しており、原核細胞の細胞質内に蓄積された溶質は外部環境中の総濃度を大きく上回る。 従って、細胞膜の内側にかかる浸透圧は10-25atmに相当すると考えられる。 膜は繊細で可塑的な構造であるため、引張強度の高い多孔質で硬い材料でできた外壁で拘束される必要がある。
ムレインはペプチドグリカンの一種で、二糖類(グリカン)が短いアミノ酸(ペプチド)鎖で架橋された高分子であり、バクテリアの細胞壁に普遍的に存在する成分である。 ペプチドグリカンには多くの種類が存在する。 すべての細菌のペプチドグリカンはN-アセチルムラミン酸を含んでおり、これはムレインの決定的な構成要素である。 古細菌の細胞壁は、タンパク質、多糖類、ペプチドグリカン様分子で構成されている場合があるが、ムレインを含むことはない。
グラム陽性菌(グラム染色で紫色のクリスタルバイオレット色素を保持する菌)では、細胞壁はペプチドグリカンの数層から構成されています。 ペプチドグリカンのシートには、グラム陽性菌に特有のテイコ酸という分子群が垂直に走っている(図14)。 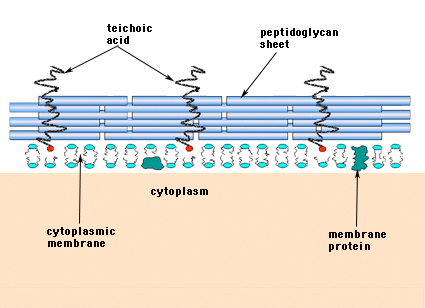
図14. グラム陽性菌の細胞壁の構造。 細胞壁は比較的厚く、ペプチドグリカンが何層にも重なっており、その間にテイコ酸がペプチドグリカンシートに直交するように存在している。
グラム陰性菌(クリスタルバイオレットを保持しない)では、細胞壁は外膜と呼ばれる膜状構造で囲まれた単層のペプチドグリカンで構成されている。グラム陰性菌の外膜には必ず、動物に対して毒性のあるユニークな成分、リポ多糖(LPSまたはエンドトキシン)を含んでいる。 グラム陰性菌の外膜は通常、細胞壁の一部と考えられている(図15)

図15. グラム陰性菌の細胞壁の構造。 細胞壁は比較的薄く、ペプチドグリカンの含有量もグラム陽性細胞壁よりはるかに少なく、また、テイコ酸も存在しない。 しかし、グラム陰性細胞壁は、ペプチドグリカン層の外側にある外膜から構成されている。
グラム陽性菌の細胞壁は厚く(15〜80ナノメートル)、何層ものペプチドグリカンで構成されている。 グラム陰性菌では、細胞壁は比較的薄く(10ナノメートル)、外膜に囲まれた1層のペプチドグリカンで構成されている。
大腸菌のペプチドグリカンの構造と配列は、すべての腸内細菌科と他の多くのグラム陰性菌の代表的なものである。 糖鎖骨格はN-アセチルグルコサミン(G)とN-アセチルムラミン酸(M)がβ1,4-グリコシド結合で交互に連結した分子である。 N-アセチルムラミン酸(M)の3位の炭素は、ピルビン酸由来のラクチルエーテル基で置換されている。 このラクチルエーテルは、L-アラニン(L-ala)、D-グルタミン酸(D-glu)、ジアミノピメリン酸(DAP)、D-アラニン(D-ala)を含むペプチド側鎖と糖鎖バックボーンをつないでいます。 MurNAcはD-glu、DAP、D-alaと同様、細菌細胞壁に特有のものである。 大腸菌のムラミン酸サブユニットを図16に示す。 大腸菌のペプチドグリカンのムラミン酸サブユニットの構造。これはほとんどのグラム陰性菌に見られるタイプのムレインである。 糖鎖骨格は、N-アセチルグルコサミン(G)とN-アセチルムラミン酸(M)の2つのアミノ糖の繰り返し重合体であり、N-アセチルグルコサミン(G)とN-アセチルムラミン酸(M)の2つの糖からなる。 N-アセチルムラミン酸にはL-ala-D-glu-DAP-D-alaからなるテトラペプチドが付加している。 c. 近接するテトラペプチド側鎖は一方のDAPと他方のD-alaとの間のペプチド間結合で互いに結合している。 d. 分子の重合体
ムレイン鎖は約100個のムラム酸サブユニットを超生物内で集合させる。 その近接性が許す限り、グリカン骨格から突出するテトラペプチド鎖は、DAP上の遊離アミノ基と近くのD-ala上の遊離カルボキシ基との間のペプチド間結合によって架橋されうる。ペプチドグリカンの細胞膜外への集合は、ペリプラズム周囲の酵素群、すなわちトランスグリコシラーゼ、トランスペプチダーゼおよびカルボキシペプチダーゼによって行われる。 ペニシリンをはじめとするβ-ラクタム系抗生物質の作用機序は、ムレイン細胞壁を形成するトランスペプチダーゼおよびカルボキシペプチダーゼ酵素を阻害することである。
ペプチドグリカン分子の糖鎖骨格は、動物血清、組織、分泌物、および食細胞系リソソームに存在するリゾチームと呼ばれる酵素によって切断されることがある。 リゾチームの機能は、細菌性病原体に対する構成的な防御として、細菌細胞を溶解することである。 涙液は4万分の1に薄めても、特定の細菌細胞を溶解する能力を保持する。 グラム陰性菌は、ペプチドグリカンが外膜に覆われているため、リゾチームによる攻撃には弱い。 リゾチームによる正確な切断部位はN-アセチルムラミン酸(M)とN-アセチルグルコサミン(G)の間のβ1,4結合であり、図16(a)に示すムラミン酸サブユニットがリゾチームが細菌のペプチドグリカンに作用した結果である<672><5552>グラム陽性菌ではペプチドグリカン中に多数の異なるペプチド配列を持っている。 最もよく研究されているのは、下の図17に示す黄色ブドウ球菌のテューリンである。 大腸菌の)DAPの代わりにジアミノ酸であるL-リジン(L-リス)があり、(グラム陰性菌の)ペプチド間結合の代わりに、リジン上の遊離アミノ基と近くのテトラペプチド側鎖のD-アラ上の遊離カルボキシ基を結ぶアミノ酸のペプチド間ブリッジが存在する。 この配置により、近接するテトラペプチド側鎖間の交差結合がより頻繁に起こるようになったと思われる。 S. aureusでは、ペプチド間橋は5つのグリシン分子からなるペプチドである(ペンタグリシン橋と呼ばれる)。 グラム陽性ムレインにおけるペプチド間結合は、グラム陰性ムレインにおけるペプチド間結合と同様に、βラクタム系抗生物質によって阻害される。 グラム陽性菌はペプチドグリカンが外膜で保護されていないため、グラム陰性菌よりもペニシリンに対する感受性が高い。 グラム陽性菌のペプチドグリカンは、テトラペプチドの3位のDAPまたはL-lysに代わるアミノ酸、およびペプチド間のブリッジの正確な組成が異なる場合がある。 グラム陽性菌には少なくとも8種類のペプチドグリカンが存在する
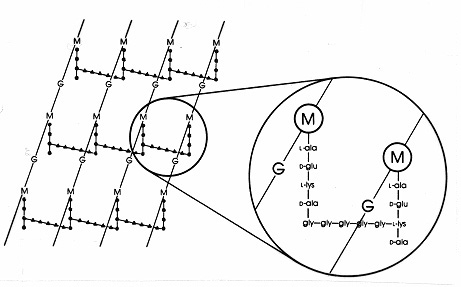
Figure 17. 黄色ブドウ球菌のペプチドグリカンシートの模式図。 G=N-アセチルグルコサミン;M=N-アセチルムラム酸;L-ala=L-アラニン;D-ala=D-アラニン;D-glu=D-グルタミン酸;L-Lys=L-リジンである。 グラム陽性菌に含まれるムレインの1種である。 L-リジンの遊離アミノ基はグリシンペンタペプチド(gly-gly-gly-gly-)に置換され、これが隣接するテトラペプチド側鎖のD-アラ由来のカルボキシ基と結合してペプチド間架橋となる。
グラム陰性菌は細胞壁にムレインの単分子層を持つが、グラム陽性菌はペプチドグリカンの数層または「ラップ」構造を持つと考えられている。 グラム陽性菌のペプチドグリカン層には、テイコ酸という一群の分子が密接に関連している。 テイコ酸は、ポリグリセロールまたはポリリビトールの直鎖ポリマーで、リン酸塩と少数のアミノ酸および糖で置換されている。 テイコ酸ポリマーは、時折、細胞膜に固定され(リポテイク酸、LTAと呼ばれる)、ペプチドグリカンの層と直角に外側に向けられるように見える。 テイコ酸の機能は不明である。 グラム陽性菌が野生で生存するためには、テイコ酸が不可欠である。 1つは、複雑なペプチドグリカンのネットワークに正電荷の物質を通すために、規則正しく配置された負電荷のチャネルを提供しているという考えである。 もう一つは、テイコ酸が細胞膜の外側にあるムラミン酸ユニットの制御と集合に何らかの形で関与しているという説である。 特に連鎖球菌では、テイコ酸が細菌の組織表面への付着に関与している例がある。
章の続き
前のページ
コメントを残す