ヘビアン理論
On 12月 14, 2021 by admin評価|生物心理学|比較|認知|発達|言語|個人差|性格|哲学|社会|
方法|統計|臨床|教育|産業|専門項目|世界心理学|
認知心理学:注意 – 意思決定 – 学習 – 判断 – 記憶 – 動機 – 知覚 – 推論 – 思考 – 認知過程認知 – 概要索引
ヘビアン理論とは、シナプス前細胞がシナプス後細胞に繰り返し持続的に刺激を与えることによってシナプスの効力が増大するというシナプス可塑性の基本メカニズムを述べたものである。 1949年にドナルド・ヘブが提唱し、ヘブの法則、ヘブの仮定、細胞集合理論とも呼ばれ、次のように述べられている。…細胞Aの軸索が細胞Bを興奮させるに十分な距離にあり、繰り返しまたは持続的にその発火に関与するとき、何らかの成長過程または代謝変化が一方または両方の細胞で起こり、Bを発火させる細胞の1つとしてAの効率が高まるのである。
この理論はしばしば「一緒に発火する細胞は一緒に配線する」と要約されるが、これは神経系を単純化しすぎており、文字通りには受け取れないし、細胞の接続強度の変化に関するヘブの元の声明を正確に表しているわけでもない。 この理論は、細胞の同時活性化によってシナプスの強度が顕著に増加する、ある種の連想学習を説明するためによく引き合いに出される。 ヘブのエングラムと細胞集合説
ヘブのエングラム説は、ニューロンがどのように結合してエングラムになりうるかに関するものである。 細胞集合体の形態と機能に関するヘブの理論は次のように理解できる:
「一般的な考えは古いもので、同時に繰り返し活動する任意の2つの細胞または細胞のシステムは、一方の活動が他方の活動を促進するように「関連」する傾向があるというものである」。 (Hebb 1949, p. 70) “ある細胞が別の細胞の発火を繰り返し補助するとき、最初の細胞の軸索は2番目の細胞のソーマと接触してシナプスノブを発達させる(あるいは既に存在していればそれを拡大する)。” (Hebb 1949, p. 63)
ゴードン・オールポートは、細胞集合論とエングラムの形成におけるその役割に関して、次のように説明される自動関連性の概念に沿って、さらなるアイデアを提示している:
「もしシステムへの入力によって活動の同じパターンが繰り返し起こるなら、そのパターンを構成する一連の活動要素はますます強く相互関連するようになるであろう」。 つまり、各要素は他のすべての要素をオンにし、(負の重みで)パターンの一部を形成していない要素をオフにする傾向がある。 別の言い方をすれば、パターン全体が「自動的な関連付け」をするようになるのである。 学習された(自動関連づけされた)パターンをエングラムと呼ぶことができる。” (Hebb 1949, p. 44)
ヘッブ理論は、全体的なレベルから分析すると、エングラムはニューロンネットまたは神経ネットワークであるという従来の見解の主要な根拠となってきた。
エリック・カンデルの研究室の研究は、海産腹足類のAplysia californicaにおけるシナプスでのヘッブ学習機構の関与についての証拠を提供している。
脊椎動物の中枢神経系シナプスのヘブ型シナプス修飾機構に関する実験は、海洋無脊椎動物で研究されている比較的単純な末梢神経系シナプスの実験よりも制御がはるかに困難である。 脊椎動物の神経細胞間の長期的なシナプス変化(長期増強など)に関する研究の多くは、脳細胞を非生理的に実験的に刺激する方法を用いている。 しかし、脊椎動物の脳で研究されてきた生理学的に適切なシナプス修飾機構のいくつかは、ヘブプロセスの例であると思われる。 そのような研究の1つは、シナプスの強度の長期的な変化が、ヘブのメカニズムと非ヘブのメカニズムの両方を通じて働く生理学的に関連するシナプス活動によって誘導され得ることを示す実験結果をレビューする
原理
人工ニューロンおよび人工ニューラルネットワークの観点から、ヘブの原理はモデルニューロン間の重みをいかに変更するかの決定方法として記述することが可能である。 2つのニューロン間の重みは、2つのニューロンが同時に活性化すれば増加し、別々に活性化すれば減少する。 同時に両方正または両方負になる傾向があるノードには強い正の重みがあり、反対になる傾向があるノードには強い負の重みがある」
このオリジナルの原理は、おそらく最も単純な重み選択形式である。 このことは、コンピュータ・プログラムに比較的容易にコード化でき、ネットワークの重みを更新するのに利用できることを意味するが、ヘブ型学習の応用の数を制限することにもなっている。 現在、ヘブ型学習という言葉は、ヘブが提唱した原理を数学的に抽象化したものを指すことが一般的である。 この意味で、ヘブ型学習は、学習ノード間の重みを調整し、各重みがノード間の関係をよりよく表すようにするものである。 375>
以下はヘブ型学習を数式で説明したものである。 (ここで、
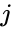
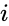
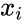
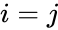
もう一つの公式的な記述としては
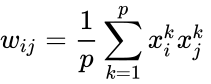
ここで、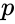

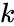
ブロッキングなどの現象や多くの神経学習現象を考慮したヘブ型学習の変形として、ハリー・クロプフの数学的モデルがある。 Klopfのモデルは非常に多くの生物学的現象を再現し、実装も簡単である。
Generalization and stability
ヘブの法則はしばしば
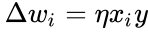
あるいは
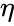
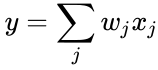
の場合で、前節の単純化では学習率も入力重みも1とするものであった。 このバージョンのルールは明らかに不安定であり、支配的な信号があるネットワークではシナプス重みは指数関数的に増減することになる。 しかし、どのようなニューロンモデルであっても、ヘブの法則は不安定であることが示される。 そのため、ニューロンのネットワークモデルでは、通常、BCM理論、Ojaの法則、一般化ヘブアルゴリズムなど、他の学習理論が採用される。
- アンチヘブ学習
- BCM理論
- 神経生物学における偶然性の検出
- デールの原理
- 一般化ヘブアルゴリズム
- リーブラ
- ロング-ヘビアンアルゴリズムも参照してください。term potentiation
- Memory
- Metaplasticity
- Neural networks
- Oja learning rule
- Tetanic stimulation
- Spike-timing-dependent plasticity
- Synaptotropic hypothesis
- Shouval, ハレル 脳の物理学 学習と記憶のシナプス基盤。 理論的アプローチ。 テキサス大学健康科学センター・ヒューストン校。 URLは2007-11-14でアクセス。
Further reading
- Hebb, D.O. (1949), The organization of behavior, New York: Wiley
- Hebb, D.O. (1961). “Distinctive features of learning in the higher animal” J. F. Delafresnaye (Ed.) Brain Mechanisms and Learning, London: オックスフォード大学出版局
- Hebb, D.O., and Penfield, W. (1940). 前頭葉の広範な両側切除後の人間の行動. 神経と精神医学のアーカイブス44:421-436.
- Allport, D.A. (1985)。 「分散記憶,モジュラーシステムと失語症」Newman, S.K. and Epstein, R. (Eds.) Current Perspectives in Dysphasia, Edinburgh: チャーチル・リビングストン.
- Bishop, C.M. (1995). パターン認識のためのニューラルネット, オックスフォード: オックスフォード大学出版局.
- Paulsen, O., Sejnowski, T. J. (2000). 活動の自然なパターンと長期的なシナプス可塑性. Current opinion in neurobiology 10 (2): 172-179.
- 概要
- Hebbian Learning tutorial(第1部:Novelty Filtering、第2部:Novelty Filtering、第3部:Hebbian Learning)。 PCA)
|
v-d-e
Neuroethology |
||
|---|---|---|
| Neurosologyにおける概念 |
Feedforward – Coincidence detector – (偶然発見器 Umwelt – 本能 – 特徴検出器 – 中心パターン発生器(CPG) -NMDA受容体 – 側方抑制 – 固定作用パターン – クローの原理-ヘブライ語理論 – 音の定位 |
200px |
| 神経倫理学の歴史 |
Theodore Holmes Bullock – Walter Heiligenberg – Niko Tinbergen- Konrad Lorenz – Eric Knudsen-Donald Griffin – Donald Kennedy – Karl von Frisch – Erich von Holst – Jörg- Neuroethologyの歴史は、1960年代後半から1960年代前半にさかのぼります。Peter Ewert |
|
| Methods in Neuroethology |
Whole Cell Patch Clamp |
|
| Model Systems in Neuroethology |
Animal Echolocation – Waggle(動物エコロケーション)-のページです。 ダンス-電気魚-ヒキガエルの視覚-カエルの聴覚の神経倫理学-ヘビの赤外線感知 |
|
このページは、Wikipedia(著者表示)からクリエイティブ・コモンズ・ライセンスのコンテンツを使用しています。
コメントを残す