サメウィーク。 メガロドンは何を食べたのか?
On 12月 13, 2021 by adminGuest Author: Jack Cooper, MSc
UoB Graduate / PhD Student, University of Swansea
これまで見てきたように、メガロドンの分類上の割り当てと最大体格に関する科学的コンセンサスには、変更や更新がありました。 しかし、その好物は、科学界で普遍的に合意されているものです。 また、珍しいことに、フィクションの中ではかなり正確に描かれているものである。 実際のクジラを食べたサメを紹介させてください!
今日の大型肉食性サメの多くは、海洋哺乳類を食べることで知られており、特に私たちの古い友人であるホオジロザメがそうです。 このサメの最も象徴的な写真のいくつかは、最新の犠牲者を顎で挟んで水面から飛び出すものである(Fig.1)。 しかし、ホホジロザメの幼魚は最初からアザラシを食べるわけではありません。 しかし、ホホジロザメの幼魚は最初からアザラシを食べているわけではなく、最初は魚や小型のサメを食べている。 パナマのガトゥン地層をもう一度見てみると、サメやエイ、耳石(魚の内耳の構造物)の化石がたくさんあり、軟体動物などの無脊椎動物もたくさんあります。 これは、メガロドンの子どもは、すぐに海洋哺乳類を狙うのではなく、魚や小型のサメも食べていたことを示しているようです。 これは、サメが成長するにつれて、好む餌を変えていくことを意味する「食性の変化(ontogenetic shift in diet)」と呼ばれるものです。 メガロドンは、成体サイズになるにつれて、より大きな餌を、おそらくはたくさん食べることで、その大きさによる代謝の要求を維持することができたはずである。

メガロドンは、その巨大なサイズに見合うだけの食料が必要だったことを考えると、ほぼ確実に日和見的な摂食をしていたのでしょう。 化石記録は、ここで私たちの最良のガイドです。 これは、生物活動の地質学的記録である痕跡化石が存在するためです。 痕跡化石には、恐竜の足跡や骨の傷など、その骨の持ち主であった動物が何かに噛まれたことを示すものがある。
さまざまな鯨類の化石から、噛み痕のあるものがいくつか見つかっている。 よく知られている例は、メリーランド州のヨークタウン地層から発見されたもので、いくつかのクジラの骨には、明らかに大きな鋸歯で作られた噛み跡がありました。 実際、メガロドンの歯がこれらの骨と直接結びついて見つかることもある。
サメは生涯を通じて何千回も歯を失ったり交換したりしているので、激しい殺しをするときに歯を失うことが多いのだ。 これが、サメの歯が化石としてよく見られる主な理由である。 傷ついた鯨の骨のすぐそばにサメの歯があるということは、餌付けが行われていたことを意味する。 しかし、古生物学者がこのような化石が発見されたときに常に考えるべきことは、これは本当に捕食だったのか、ということです。 8769>
捕食は、ホホジロザメ(図2)やイタチザメなど、現代の大型捕食性サメに共通する摂食行動である。 これらのサメは、海面に浮かぶクジラの死骸に遭遇すると、食べ放題のビュッフェになるのです。 捕食に失敗すると、エネルギーを無駄に消費することになる。 メガロドンはその巨大な体格を維持する必要があったため、死んだクジラに遭遇してもタダ飯を見過ごすことはあまり意味がないだろう。 したがって、現代の大型のサメがどのように餌を食べるかを考えると、おそらくメガロドンはチャンスがあれば餌を漁ったのだろうと、ほとんどの人は思うだろう。

このように、メガロドンが捕食を試みたことを示唆する、特に注目すべき化石が1つあるのです。 それは、メガロドンが捕食に失敗したことを示す化石です。 2010年に発表された論文では、ノースカロライナ州のヨークタウン層(そう、2つのヨークタウン層)から出土した鯨類の肋骨の部分化石を紹介しています。 この化石にも噛み跡のような傷があったが、注目すべきは、その傷を覆うように骨が編まれているように見えたことである。 著者らはこれを、噛み痕による骨の外傷が回復する過程で感染症が発生した痕跡と解釈している。 また、この噛み痕は鋸歯状であることから、メガロドンと同様に鋸歯状であったことが示唆された。 この研究は、メガロドンまたは他の大きなサメが、この化石の動物を襲いましたが、犠牲者は生き延びたものの、6週間後にその傷から感染症で死亡したことを示唆しました。 この地は、ゴードン・ハベルが例外的なカルカロドン化石(Q1で説明)を発見した場所と同じなので、おそらく運命的なものだったのでしょう。 ここで見つかった噛み跡には、頭蓋骨や肋骨が含まれており、このサメは対象を横から噛み砕いたと考えられる。 最も重要なことは、この遺骨が初めて1つの種に特定されたことだ。その種とは、小型のヒゲクジラであるピスコバラエナ・ナナ(Piscobalaena nana)である。 他の痕跡化石と同様に、噛み跡は大きな鋸歯とよく一致しており、メガロドンが攻撃したことを示す好例である(図3)。 したがって、メガロドンは、イルカやウミガメなどの小型の獲物もターゲットにしていた可能性が高いという指摘と非常に一致し、小型のクジラも(捕食または清掃のために)ターゲットにしていたように思われました。 Alberto Gennariによるアートワーク。 画像はCollareta et al. 2017 .化石から捕食戦略を解釈することは難しいが、現代のサメはメガロドンがどのように狩りを行っていたかを教えてくれるかもしれない。 その咬合力は、鯨の骨を損傷するために例外的に強力だったでしょう。 ホホジロザメの頭蓋骨のコンピューターモデルから、その咬合力は約1.8トンを超え、生きている動物の中で最も強い咬合力の一つであることが提唱された。 この研究は、メガロドンの咬合力がさらに大きく、108,514 ~ 182,201 Nm と推定されることを示唆しています。 サメはバーストスピードと呼ばれる、獲物を捕らえるときに使われる高速の泳ぎをすることがあります。 しかし、通常、サメが何気なく泳いでいるときのスピードは、いわゆる持続泳速度である。 これは種によって異なるが、David Jacobyが率いるある研究では、64の先行研究から得られた26種のデータを組み合わせ、持続的遊泳速度とサメの代謝や体格との間のスケーリング関係を予測するモデルを開発した。 代謝を組み込んだそのモデルでは、次の式で要約される 0.173 というスケーリング指数が明らかになりました。
Swim Speed (ms ¯¹) ∝ Mass (kg)^0.173
ところが、データを系統的に補正すると、最小泳速度が体重に対して 0.15 というやや低い指数で比例することが判明したのです。 上記の式はさらに、比率を合わせるために定数1を仮定しているが、この定数は実際には0.266であることがデータから判明した。 そこで、泳ぐ速度(ms¹)=0.266質量(kg)^0.15
ここから、質量に応じたメガロドンの持続泳速を算出することができる。 例えば、ゴットフリードが15.9mのサメの質量として計算した48,000kgのメガロドンは、1.34m/s(時速約4.8km)で連続的に泳いでいることがわかる。 これを52,000kgのメガロドンに当てはめると、1.36m/s、時速4.9kmで泳ぎ続けることになる。一方、極端な話、10万kgのサメをシミュレーションすると、このモデルは1.50m/s、時速5.4kmの泳ぎになるのだ。 このモデルは、メガロドンの泳ぎ速度の計算だけでなく、複数のサメの種に対するモデルとしても非常に有効であることが証明されました(図 4)。
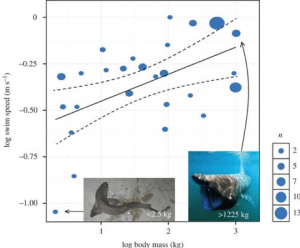
より引用:しかし、サメが獲物を素早くさらうために使うのはバーストスピードである。 ホホジロザメはこれを利用して、下から獲物に突進して不意を突く「ブリーチング行動」をとる(図1). このような行動ができるのは、逆光によってうまくカモフラージュしているからである。 つまり、メガロドンもカウンターシェーディングをしていた可能性が高く、生態や体の構造もホホジロザメと似ているので、メガロドンもブリーチングが可能だったのだろうかという疑問が湧いてきます。 ホホジロザメの破裂速度は、中温性という生理的適応によってもたらされる。 これは最近メガロドンも持っていたことが示唆されています(詳しくはQ5で説明します)。 中温性魚類は一般に外温性魚類よりも持続的な泳ぎと破裂の速度が速く、これを利用して、メガロドンの破裂速度は時速 37.15 km であった可能性が指摘されている
このように大きなサメとしては信じられないほど速いスピードは、突破するには十分であろう。 このほか、現在 2 番目に大きいサメであるウバザメ (Cetorhinus maximus) も、ブリーチングが観察されています。 おそらく、巨大なメガロドンは、最新の食料を手に入れるために、本当に水から飛び出すことができたのだろう。 まだわかりませんが、私の個人的な、そして(やや専門的な)意見としては、それは十分に可能であったかもしれません。
このように幅広いメニューを持つメガロドンは、頂点捕食者で、自分自身の捕食者は知られていません。 このことは、2つの重要なことを意味します。 まず、獲物をめぐって激しい競争を繰り広げ、それが結果的に絶滅の一因となった可能性があります。 第二に、頂点捕食者であるメガロドンは、獲物の個体数を抑制し、健全な生態系を維持するため、その生態系にとって最も重要な存在であっただろう。 頂点捕食者であるサメの損失は、不可逆的な連鎖的影響をもたらし、後に残るコミュニティを変化させる。 メガロドンがいなくなったからこそ、海洋生態系が成り立っている部分もある。 そして、現代の海の巨大なクジラたちは、その結果、安心して過ごすことができるのです。
- Estrada JA, Rice AN, Natanson LJ & Skomal GB 2006. を用いた、ホホジロザメの個体発生的な摂餌生態の復元。 Ecology 87, 829-834.
- Pimiento C, Ehret DJ, MacFadden BJ & Hubbell G 2010. パナマの中新世から得られた絶滅した巨大鮫メガロドンの古代の生育地。 PLoS One 5, e10552.
- Pimiento C, González-Barba G, Ehret DJ, Hendy AJ, MacFadden BJ & Jaramillo C 2013. パナマの後期中新世ガトゥン層から出土したサメとエイ(Chondrichthyes, Elasmobranchii). J. Paleontol. 87, 755-774.
- Purdy RW 1996. 化石ホオジロザメの古生態学. In: また,”萌え “と “萌え “を融合させた “萌え “研究にも取り組んでいる. 鮮新世カリブ海パラグアナ層から出土したオオテンジクザメとクジラ類の栄養学的相互作用。 Paläontol. Z. 82, 204-208.
- Fallows C, Gallagher A. & Hammerschlag N 2013. ホワイト・シャーク(Carcharodon carcharias)がクジラを捕食する様子と、頂点の捕食者の生態をさらに形成する潜在的な役割。 PLoS One, 8, e60797.
- Dudley SFJ, Anderson-Reade MD, Thompson GS & McMullen PB 2000.鯨を捕食するホオジロザメ(Carcharodonarcharias)。 鯨の死骸をホホジロザメ(Carcharodon carcharias)とイタチザメ(Galeocerdo cuvier)が同時に漁獲したこと。 Fish. Bull. 98, 646-649.
- Kallal RJ, Godfrey SJ & Ortner DJ 2010. 鮮新世鯨類の肋骨の骨反応から捕食イベントの短期生存を示す。 Int. J. Osteoarchaeol. 22, 253-260.
- Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M & Bianucci G 2017. 絶滅した巨大サメCarcharocles megalodonは小さな獲物を狙っていたのか? ペルーの後期中新世から出土した海洋哺乳類遺体の咬傷痕。 Palaeogeogr. Palaeoclimatol. Palaeoecol. 469, 84-91.
- Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, Ferrara TL, Cunningham E, Dean MN & Summers AP 2008. ホオジロザメの顎の力学の3次元コンピュータ解析:ホオジロザメはどのくらい強く噛むことができるのか? J. Zool. 276, 336-342.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2015. サメの泳ぎ速度のスケーリングは代謝によるものか? Biol. Lett. 11, 20150781.
- Gottfried MD, Compagno LJV & Bowman SC 1996. 巨大な “メガトース “サメCarcharodon megalodonのサイズと骨格解剖学。 In: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 55-66, Academic Press, San Diego.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2016. サメの泳ぎ速度のスケーリングは代謝によって駆動されるのか」の訂正。 Biol. Lett. 12, 20160775.
- Watanabe YY, Payne NL, Semmens JM, Fox A & Huveneers C 2019.を参照。 内熱性ホオジロザメの採餌時の遊泳戦略とエネルギッシュな行動。 J. Exp. Biol. 222, jeb185603.
- Cooper JA, Pimiento C, Ferrón HG & Benton MJ (in press). 絶滅した巨大ザメOtodus megalodonの体寸法。 2次元復元。 Sci. Rep.
- Watanabe YY, Goldman KJ, Caselle JE, Chapman DD & Papastamatiou YP 2015. アニマルトラッキングデータの比較解析により、魚類における内温性の生態学的意義が明らかになった。 Proc. Nat. Acad. Sci. 112, 6104-6109.
- Ferrón HG 2017. Some extinct macropredatory sharksにおけるgigantismのトリガーとしてのregional endothermy。 PLoS One, 12, e0185185.
- Johnston EM, Halsey LG, Payne NL, Kock AA, Iosilevskii G, Whelan B & Houghton JD 2018. 例外的なブリーチングイベントによって明らかになったウバザメの潜在的な力。 Biol. Lett. 14, 20180537.
- Myers RA, Baum JK, Shepherd TD, Powers SP & Peterson CH 2007. 沿岸海域からの頂点捕食性サメの喪失によるカスケード効果. Science, 315, 1846-1850.
Edited by Rhys Charles
.
コメントを残す