Struttura e funzione delle cellule batteriche
Il Ottobre 10, 2021 da adminStruttura e funzione delle cellule batteriche (pagina 5)
(Questo capitolo ha 10 pagine)
© Kenneth Todar, PhD
Parete cellulare
Le pareti cellulari dei batteri meritano un’attenzione speciale per diverse ragioni:
1. Sono una struttura essenziale per la vitalità, come descritto sopra.
2. Sono composte da componenti unici che non si trovano da nessun’altra parte in natura.
3. Sono uno dei siti più importanti per l’attacco degli antibiotici.
4. Forniscono ligandi per l’aderenza e recettori per farmaci o virus.
5. Causano sintomi di malattia negli animali.
6. Forniscono distinzione immunologica e variazione immunologica tra i ceppi di batteri.
La maggior parte dei procarioti hanno una parete cellulare rigida. La parete cellulare è una struttura essenziale che protegge il cellprotoplasto dai danni meccanici e dalla rottura osmotica o lisi.I procarioti di solito vivono in ambienti relativamente diluiti in modo tale che l’accumulo di soluti all’interno del citoplasma delle cellule procariotiche supera di molto la concentrazione totale nell’ambiente esterno. Così, la pressione osmotica contro l’interno della membrana plasmatica può essere l’equivalente di 10-25atm. Poiché la membrana è una struttura delicata e plastica, deve essere trattenuta da una parete esterna fatta di materiale poroso e rigido che ha un’alta resistenza alla tensione. Tale materiale è la mureina, l’onnipresente componente delle pareti cellulari batteriche.
La mureina è un tipo unico di peptidoglicano, un polimero di disaccaridi (glicano) reticolati da brevi catene di amminoacidi (peptide). Esistono molti tipi di peptidoglicano. Tutti i peptidoglicani batterici contengono acido N-acetilmuramico, che è il componente definitivo della mureina. Le pareti cellulari degli Archaea possono essere composte da proteine, polisaccaridi o molecole simili ai peptidoglicani, ma non contengono mai mureina. Questa caratteristica distingue i batteri dagli archei.
Nei batteri Gram-positivi (quelli che trattengono il colorante viola cristallo quando vengono sottoposti alla procedura di colorazione Gram), la parete cellulare consiste di diversi strati di peptidoglicano. Perpendicolarmente ai fogli di peptidoglicano c’è un gruppo di molecole chiamate acido teico che sono uniche per la parete cellulare dei Gram-positivi (Figura 14). 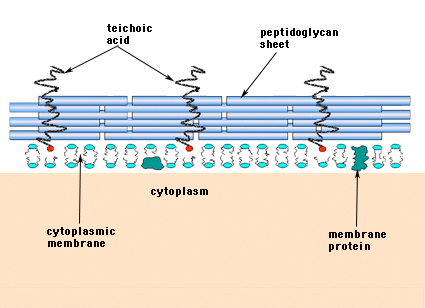
Figura 14. Struttura della parete cellulare dei batteri Gram-positivi. La parete è relativamente spessa e consiste di molti strati di peptidoglicano intervallati da acidi teicoici che corrono perpendicolarmente ai fogli di peptidoglicano.
Nei batteri Gram-negativi (che non trattengono il cristalvioletto), la parete cellulare è composta da un singolo strato di peptidoglicani circondato da una struttura membranosa chiamata membrana esterna. La membrana esterna dei batteri Gram-negativi contiene invariabilmente un unico componente, lipopolisaccaride (LPS o endotossina), che è tossico per gli animali. Nei batteri Gram-negativi la membrana esterna è solitamente considerata come parte della parete cellulare (Figura 15).

Figura 15. Struttura della parete cellulare dei Gram-negativi. La parete è relativamente sottile e contiene molto meno peptidoglicano della parete dei Gram-positivi; inoltre, gli acidi teicoici sono assenti. Tuttavia, la parete cellulare Gram negativa è costituita da una membrana esterna che si trova al di fuori dello strato di peptidoglicano. La membrana esterna è attaccata al foglio di peptidoglicano da un gruppo unico di molecole lipoproteiche.
Nei batteri Gram-positivi, la parete cellulare è spessa (15-80nanometri), costituita da diversi strati di peptidoglicano. Nei batteri Gram-negativi la parete cellulare è relativamente sottile (10 nanometri) ed è composta da un singolo strato di peptidoglicani circondato da una membrana esterna.
La struttura e la disposizione dei peptidoglicani in E.coli è rappresentativa di tutte le Enterobacteriaceae, così come di molti altri batteri Gram-negativi. La spina dorsale del glicano è costituita da molecole alternate di N-acetilglucosamina (G) e acido N-acetilmuramico (M) collegate da un legame beta 1,4-glicoside. Il 3-carbonio dell’acido N-acetilmuramico (M) è sostituito da un gruppo lattile etere derivato dal piruvato. Il lattiletere collega la spina dorsale del glicano a una catena laterale peptidica che contiene L-alanina, (L-ala), D-glutammato (D-glu), acido diammino-pimelico (DAP), e D-alanina (D-ala). MurNAc è unico per le pareti cellulari batteriche, così comeD-glu, DAP e D-ala. La subunità dell’acido muramico di E. coli è mostrata nella figura 16 qui sotto.

Figura 16. La struttura della subunità dell’acido muramico del peptidoglicano di Escherichia coli. Questo è il tipo di mureina che si trova nella maggior parte dei batteri Gram-negativi. La spina dorsale del glicano è un polimero ripetuto di due amino zuccheri, N-acetilglucosamina (G) e acido N-acetilmuramico (M). Attaccato all’acido N-acetilmuramico è un atetrapeptide costituito da L-ala-D-glu-DAP-D-ala. b. Struttura abbreviata della subunità di acido muramico. c. Le catene laterali tetrapeptidiche vicine possono essere legate l’una all’altra da un legame interpeptidico tra DAP su una catena e D-ala sull’altra. d. La forma polimerica della molecola.
Filamenti di mureinare assemblati nel periplasma da circa 10 subunità di acido muramico. Dove la loro vicinanza lo permette, le catene tetrapeptidiche che sporgono dalla spina dorsale del glicano possono essere reticolate da un legame interpeptidico tra un gruppo amminico libero sul DAP e un gruppo carbossilico libero su un D-ala vicino.L’assemblaggio del peptidoglicano all’esterno della membrana plasmatica è mediato da un gruppo di enzimi periplasmatici, che sono transglicosilasi, transpeptidasi e carbossipeptidasi. Il meccanismo d’azione della penicillina e degli antibiotici beta-lattamici correlati è quello di bloccare gli enzimi transpeptidasi e carbossipeptidasi durante il loro assemblaggio della parete cellulare murenica. Quindi, gli antibiotici beta-lattamici sono detti “bloccare la sintesi della parete cellulare” nei batteri.
La spina dorsale glicanica della molecola peptidoglicano può essere scissa da un enzima chiamato lisozima che è presente nel siero animale, nei tessuti e nelle secrezioni, e nel lisosoma fagocitico. La funzione del lisozima è quella di sciogliere le cellule batteriche come difesa costitutiva contro gli agenti patogeni batterici. Alcuni batteri Gram-positivi sono molto sensibili al lisozima e l’enzima è abbastanza attivo a basse concentrazioni. Le secrezioni lacrimali (lacrime) possono essere diluite 1:40.000 e conservano la capacità di lisciare alcune cellule batteriche. I batteri Gram-negativi sono meno vulnerabili all’attacco del lisozima perché il loro peptidoglicano è protetto dalla membrana esterna. Il sito esatto della scissione lisozimica è il legame beta 1,4 tra l’acido N-acetilmuramico (M) e la N-acetilglucosamina (G), in modo tale che la subunità di acidouramico mostrata nella Figura 16(a) è il risultato dell’azione del lisozima sul peptidoglicano batterico.
Nei batteri Gram-positivi ci sono numerose disposizioni diverse di peptidi tra i peptidoglicani. Il più studiato è quello dello Staphylococcus aureus mostrato nella figura 17 qui sotto. Al posto del DAP (in E. coli) c’è il diaminoacido, L-lisina (L-lis), e al posto del legame interpeptidico (nei Gram-negativi) c’è un ponte interpeptidico di aminoacidi che collega un gruppo amminico libero sulla lisina a un gruppo carbossilico libero su D-ala di una vicina catena laterale tetrapeptidica. Questa disposizione permette apparentemente un legame incrociato più frequente tra le catene laterali tetrapeptidiche vicine. In S. aureus, il ponte interpeptidico è un peptide composto da 5 molecole di glicina (chiamato ponte di pentaglicina). L’assemblaggio del ponte interpeptidico nella mureina Gram-positiva è inibito dagli antibiotici beta-lattamici allo stesso modo del legame interpeptidico nella mureina Gram-negativa. I batteri Gram-positivi sono più sensibili ai batteri topenicillinthan Gram-negativi perché il peptidoglicano non è protetto da una membrana esterna ed è una molecola più abbondante. Nei batteri Gram-positivi, i peptidoglicani possono variare nell’amminoacido al posto del DAP o L-lys in posizione 3 del tetrapeptide, e nell’esatta composizione del ponte interpeptidico. Nei batteri Gram-positivi esistono almeno otto diversi tipi di peptidoglicano.
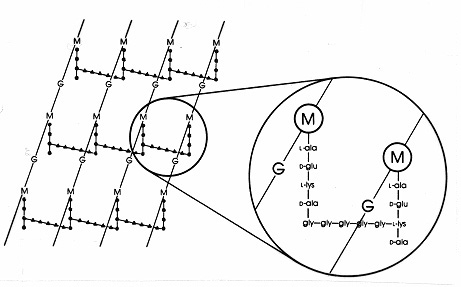
Figura 17. Diagramma schematico del foglio di peptidoglicano di Staphylococcus aureus. G = N-acetil-glucosamina; M = acido N-acetil-muramico; L-ala = L-alanina; D-ala = D-alanina; D-glu= acido D-glutammico; L-lys = L-lisina. Questo è un tipo di mureina che si trova nei batteri Gram-positivi. Rispetto al peptidoglicano di E. coli (Figura 7) c’è L-lys al posto di DAP (acido diaminopimelico) in thetetrapeptide.The gruppo amminico libero di L-lys è sostituito con un glicinepentapeptide(gly-gly-gly-gly-gly-) che poi diventa un collegamento interpeptide bridgeforminga con un gruppo carbossilico da D-ala in un tetrapeptide adiacente catena laterale. I peptidoglicani Gram-positivi differiscono da specie a specie, principalmente per quanto riguarda gli aminoacidi nella terza posizione della catena tetrapeptidica e nella composizione aminoacidica del ponte interpeptidico.
I batteri Gram-negativi possono contenere un singolo strato monomolecolare di mureina nelle loro pareti cellulari mentre i batteri Gram-positivi si pensa abbiano diversi strati o “involucri” di peptidoglicano. Strettamente associati agli strati di peptidoglicano nei batteri Gram-positivi sono un gruppo di molecole chiamate acidi teicoici. Gli acidi teicoici sono polimeri lineari di poliglicerolo o poliribitolo sostituiti con fosfati e alcuni amminoacidi e zuccheri. I polimeri di acido teico sono occasionalmente ancorati alla membrana plasmatica (chiamati acido lipoteico, LTA) apparentemente diretti verso l’esterno ad angolo retto rispetto agli strati di peptidoglicano. Le funzioni dell’acido teico non sono note. Sono essenziali per la vitalità dei batteri Gram-positivi in natura. Un’idea è che essi forniscono un canale di cariche negative orientate regolarmente per infilare sostanze caricate positivamente attraverso la complicata rete del peptidoglicano. Un’altra teoria è che gli acidi teicoici siano in qualche modo coinvolti nella regolazione e nell’assemblaggio delle subunità di acidi muramici all’esterno della membrana plasmatica. Ci sono casi, in particolare negli streptococchi, in cui gli acidi teicoici sono stati implicati nell’aderenza dei batteri alle superfici dei tessuti.
capitolo continuato
Pagina precedente
Lascia un commento