Nucleo olivario superiore
Il Novembre 12, 2021 da admin| Cervello: Complesso olivario superiore | ||
|---|---|---|
 |
||
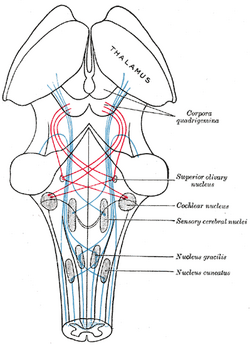
| Schema che mostra il corso delle fibre del lemnisco; lemnisco mediale in blu, laterale in rosso. (Il nucleo olivario superiore è etichettato al centro a destra.) | ||
| ] | ||
| Latino | nucleo olivare superiore | |
| di Gray | soggetto #187 787 | |
| parte di | ||
| Componenti | ||
| Arteria | ||
| Vein | ||
| BrainInfo/UW | {{BrainInfoType}}-565 | |
| MeSH | ||
Il complesso olivario superiore (o SOC o oliva superiore) è una piccola massa di sostanza grigia situata sulla superficie dorsale della parte laterale del corpo trapezoidale. Piccolo nell’uomo, ma ben sviluppato in certi animali, presenta una struttura simile a quella del nucleo olivare inferiore, ed è situato immediatamente sopra di esso. Alcune delle fibre del corpo trapezoidale terminano arborizzandosi intorno alle cellule di questo nucleo, mentre altre nascono da queste cellule.
Fisiologia
Il nucleo olivare superiore svolge una serie di ruoli nell’udito. L’oliva superiore mediale (MSO) è un nucleo specializzato che si ritiene misuri la differenza temporale di arrivo dei suoni tra le orecchie (la differenza temporale interaurale o ITD). L’ITD è uno spunto importante per determinare l’azimut dei suoni a bassa frequenza, cioè per localizzarli sul piano azimutale – il loro grado a sinistra o a destra.
L’oliva laterale superiore (LSO) si ritiene essere coinvolto nella misurazione della differenza di livello dei suoni tra le orecchie (la differenza di livello interaurale o ILD). L’ILD è un secondo importante spunto nel determinare l’azimut dei suoni ad alta frequenza.
Relazione con il sistema uditivo
Il complesso olivario superiore è generalmente situato nel ponte, ma nell’uomo si estende dal midollo allungato rostrale al mid-pon e riceve proiezioni prevalentemente dal nucleo cocleare anteroventrale attraverso la stria acustica ventrale, anche se il nucleo posteroventrale proietta anche lì attraverso la stria acustica intermedia. È in questo sito che si verificano le prime grandi interazioni binaurali.
Nuclei primari
Il complesso olivario superiore è diviso in tre nuclei primari, il MSO, LSO e il nucleo mediale del corpo trapezoidale, e diversi nuclei periolivari più piccoli. Questi tre nuclei sono i più studiati e quindi meglio compresi. Tipicamente, sono considerati come formanti la via ascendente di localizzazione azimutale.
Olivo superiore mediale (MSO)
Si pensa che l’olivo superiore mediale aiuti a localizzare l’azimut di un suono, cioè l’angolo a sinistra o a destra dove si trova la sorgente sonora. Il primo istinto potrebbe essere quello di pensare che questo nucleo includa informazioni verticali, ma non è così; le informazioni elaborate nel MSO sono limitate al piano orizzontale. Le cellule fusiformi non proiettano a nulla a livello del ponte, ed entrano in gioco solo a livello del collicolo inferiore. Sono presenti solo dati orizzontali, ma provengono da due diverse fonti auricolari, il che aiuta la localizzazione del suono sull’asse azimutale. Il modo in cui l’oliva superiore fa questo è misurando le differenze di tempo tra i segnali di due orecchie che registrano lo stesso stimolo. Il viaggio intorno alla testa richiede circa 700 μs, e si presume che l’oliva superiore mediale sia in grado di rilevarlo. Infatti, si osserva che le persone possono rilevare differenze interaurali fino a 10 microsecondi. Il nucleo è organizzato tonotopicamente, ma la proiezione azimutale del campo recettivo è “molto probabilmente una mappa complessa e non lineare”.
Le proiezioni dell’oliva superiore mediale terminano densamente nel nucleo centrale omolaterale del collicolo inferiore. La maggior parte di questi assoni sono considerati di “forma rotonda” o di tipo R. Questi assoni R sono per lo più glutamatergici e contengono vescicole sinaptiche rotonde e formano giunzioni sinaptiche asimmetriche.
- Questo è il più grande dei nuclei e nell’uomo contiene circa 15.500 neuroni.
- Ogni MSO riceve input bilaterali a bassa frequenza dalle AVCNs destra e sinistra.
- L’uscita è al lemnisco laterale omolaterale e infine al collicolo inferiore.
- L’MSO risponde meglio a stimoli binaurali.
- La sua funzione principale comporta il rilevamento delle differenze nel tempo di arrivo dei suoni alle due orecchie che fa parte del processo di localizzazione (ITD).
Oliva laterale superiore (LSO)
Questa oliva ha funzioni simili all’oliva mediale superiore, ma utilizza l’intensità per localizzare la sorgente sonora. Questa è la parte del tronco cerebrale che etichetta il suono più forte dall’orecchio sinistro come proveniente dal lato sinistro. L’oliva laterale riceve input da entrambi i nuclei cocleari, dalle cellule cespugliose sferiche AVCN omolaterali e dalle cellule cespugliose globulari AVCN controlaterali attraverso il MNTB omolaterale. MNTB cellule principali sono glicinergici, fornendo una fonte di inibizione al LSO, e formando la base della sensibilità ILD. Proiezioni da entrambi i nuclei cocleari sono principalmente ad alta frequenza, e queste frequenze sono successivamente rappresentato dalla maggior parte dei neuroni LSO (>2/3 oltre 2-3kHz nel gatto). Ulteriori ingressi derivano dal LNTB omolaterale (glicinergico, vedi sotto), che forniscono informazioni inibitorie dal nucleo cocleare omolaterale. Un altro possibile input inibitorio deriva dalle cellule non sferiche AVCN omolaterali. Queste cellule sono globulari cespugliose o multipolari (stellate). Uno di questi due ingressi potrebbe fornire la base per l’inibizione ipsilaterale visto nelle mappe di risposta che fiancheggiano l’eccitazione primaria, affilando sintonia frequenza dell’unità.
I progetti LSO bilateralmente al nucleo centrale del collicolo inferiore (ICC). Proiezioni omolaterali sono principalmente inibitorio (glicinergico), e controlaterale eccitatorio. Ulteriori obiettivi di proiezione includono i nuclei dorsale e ventrale del Lemnisco laterale (DNLL & VNLL). Le proiezioni GABAergiche dal DNLL formano la fonte primaria di GABA nel tronco encefalico uditivo, e proiettano bilateralmente all’ICC e al DNLL controlaterale. Queste connessioni convergenti eccitatorie e inibitorie possono agire per diminuire la dipendenza dal livello della sensibilità ILD nel ICC rispetto al LSO.
Proiezioni aggiuntive formano il Bundle Olivocochlear laterale (LOC), che innerva le cellule ciliate interne cocleari. Si pensa che queste proiezioni abbiano una lunga costante di tempo, e agiscono per normalizzare il livello sonoro rilevato da ciascun orecchio per aiutare la localizzazione del suono. Esistono notevoli differenze di specie: I neuroni di proiezione LOC sono distribuiti all’interno del LSO nei roditori, e circondano il LSO nei predatori (es. gatto).
Nucleo Mediale del Corpo Trapezoidale (MNTB)
- Il MNTB è la parte più piccola del SOC; la presenza del MNTB nel tronco encefalico umano è in dubbio.
- Ogni neurone riceve solo input ad alta frequenza attraverso un calice di tenuta dal AVCN controlaterale, e la sua uscita proietta al LSO omolaterale.
- Ci sono due tipi di risposta trovati: un ‘tipo chopper’ simile alle cellule del fuso nel AVCN e un tipo primario che è simile a quelle delle cellule cespugliose nel AVCN.
Nuclei periolivari
Il SOC è composto da tra sei e nove nuclei periolivari, a seconda del ricercatore citato, in genere nominato in base alla loro posizione rispetto ai nuclei primari. Questi nuclei circondano ciascuno dei nuclei primari e contribuiscono al sistema uditivo ascendente e discendente. Questi nuclei formano anche la fonte del fascio olivococcleare, che innerva la coclea. Nella cavia, le proiezioni ascendenti ai collicoli inferiori sono principalmente ipsilaterali (>80%), con la più grande fonte singola proveniente dallo SPON. Inoltre, i nuclei ventrali (RPO, VMPO, AVPO, & VNTB) sono quasi interamente ipsilaterale, mentre i restanti nuclei progetto bilateralmente.
| Nome | Cat | Guinea Pig | Rat | Mouse | ||
|---|---|---|---|---|---|---|
| LSO | X | X | X | X | ||
| MSO | X | X | X | X | ||
| MNTB | X | X | X | X | X | |
| LNTB | X | X | “LVPO” | X | ||
| ALPO | X | X | ||||
| PVPO | X | X | ||||
| PPO | X | X | “CPO” | |||
| VLPO | X | |||||
| DPO | X | X | X | |||
| DLPO | X | X | X | |||
| VTB | X | X | “MVPO” | X | ||
| AVPO | X | |||||
| VMPO | X | X | ||||
| RPO | X | X | X | |||
| SPN | “DMPO” | X | X | X |
Nucleo Ventrale del Corpo Trapezoidale (VNTB)
- Il VNTB è un piccolo nucleo situato lateralmente al MNTB e ventrale al MSO.
- Composto da una popolazione eterogenea di cellule, questo nucleo proietta a molti nuclei uditivi, e forma il fascio olivocleare mediale (MOC) che innerva le cellule ciliate esterne cocleari. Queste cellule contengono fibre elettromotili, e agiscono come amplificatori/attenuatori meccanici all’interno della coclea.
- Il nucleo proietta a entrambi i CI, senza cellule che proiettano bilateralmente.
Nucleo laterale del corpo trapezoidale (LNTB)
- Situato ventralmente al LSO
- Le cellule cespugliose sferiche AVCN proiettano collaterali bilateralmente, e le cellule cespugliose globulari proiettano collaterali ipsilateralmente ai neuroni LNTB.
- Le cellule sono immunoreattive per la glicina, e sono etichettate retrogradamente dopo l’iniezione di glicina triziata nel LSO
- Il nucleo proietta ad entrambi IC, con poche cellule che proiettano bilateralmente, così come il LSO ipsilaterale.
- Grandi cellule multipolari proiettano al nucleo cocleare, ma non il IC, sia nel gatto che nella cavia.
- Gli ingressi avvengono spesso attraverso i bulbi terminali di tenuta, producendo una trasduzione del segnale molto veloce.
Nucleo Periolivario Superiore (SPON) (Nucleo Periolivario Dorsomediale (DMPO))
- Situato direttamente dorsale al MNTB
- Nel ratto, lo SPON è un nucleo GABAergico omogeneo. Questi neuroni organizzati tonotopicamente ricevono input eccitatori dal polipo e dalle cellule multipolari nel nucleo cocleare ventrale controlaterale, un input glicinergico (inibitorio) dal MNTB omolaterale, un input GABAergico (inibitorio) sconosciuto, e proiettano al ICC omolaterale. La maggior parte dei neuroni rispondono solo all’offset di uno stimolo, può bloccare la fase a stimoli AM fino a 200 Hz, e può costituire la base per la selettività durata ICC. In particolare, i neuroni SPON non ricevono input discendente dal IC, e non proietta alla coclea o nucleo cocleare come fanno molti nuclei periolivari.
- Nella cavia, le cellule multipolari da rotonde a ovali proiettano a entrambi gli IC, con molte cellule che proiettano bilateralmente. Le cellule più allungate che proiettano al nucleo cocleare non proiettano all’ICC. Sembra che ci siano due popolazioni di cellule, una che proietta ipsilateralmente e una che proietta bilateralmente.
- La maggior parte delle informazioni erano venute da SPON roditori, a causa delle dimensioni prominenti del nucleo in queste specie, con pochissimi studi sono stati fatti in gatto DMPO, nessuno dei quali era esteso.
Nucleo Periolivario Dorsale (DPO)
- Situato dorsalmente e medialmente al LSO
- Contiene sia unità EE (eccitate da entrambe le orecchie) che E0 (eccitate solo dall’orecchio controlaterale).
- I neuroni sono organizzati tonotopicamente, e ad alta frequenza.
- Può appartenere ad un unico nucleo insieme al DLPO
- Il nucleo proietta ad entrambi gli IC, senza cellule che proiettino bilateralmente.
Nucleo Periolivario Dorsolaterale (DLPO)
- Situato dorsalmente e lateralmente al LSO
- Contiene sia unità EE (eccitate da entrambe le orecchie) che E0 (eccitate solo dall’orecchio controlaterale).
- I neuroni sono organizzati tonotopicamente, e a bassa frequenza.
- Può appartenere ad un unico nucleo insieme al DPO
- Il nucleo proietta ad entrambi gli IC, con poche cellule che proiettano bilateralmente.
Nucleo Periolivario Ventrolaterale (VLPO)
- Situato ventralmente e all’interno della collina ventrale del LSO
- Contiene sia unità EI (eccitate dall’orecchio controlaterale e inibite da quello omolaterale) che E0 (eccitate solo dall’orecchio controlaterale).
- I neuroni sono organizzati tonotopicamente, e ad alta frequenza.
Nucleo Periolivario Anterolaterale (ALPO)
- Il nucleo proietta ad entrambi gli IC, senza cellule che proiettano bilateralmente.
- Grandi cellule multipolari proiettano al nucleo cocleare, ma non all’IC, sia nel gatto che nella cavia.
Nucleo Periolivario Ventromediale (VMPO)
- Situato tra il MSO e il MNTB.
- Manda proiezioni all’ICC bilateralmente.
- Il nucleo proietta ad entrambi gli IC, senza cellule che proiettano bilateralmente.
Nucleo Periolivario Rostrale (RPO) (Nucleo Periolivario Anteriore (APO))
- Situato tra il polo rostrale del MSO e il VNLL
Nucleo Periolivare Caudale (CPO) (Nucleo Periolivare Posteriore (PPO))
- Situato tra il polo caudale del MSO e il nucleo facciale (7N)
Nucleo Periolivare Posteroventrale (PVPO)
- Il nucleo proietta ad entrambi gli IC, senza cellule che proiettano bilateralmente.
Fisiopatologia
L’autopsia di una donna di 21 anni con autismo, epilessia e ritardo mentale ha trovato una quasi completa assenza dell’oliva superiore.
Vedi anche
- Corpo olivare
- Kulesza RJ, Cytoarchitecture of the human superior olivary complex: Medial and lateral superior olive. Hearing Research 225(2007) 80-90
- 2.0 2.1 Oliver DL, et al. Proiezioni assonali dal laterale e mediale oliva superiore al collicolo inferiore del gatto: uno studio utilizzando microscopia elettronica autoradiografia. J Comp Neurol. 1995 Sep 11;360(1):17-32 Cite errore: Invalid
<ref>tag; nome “R4” definito più volte con contenuto diverso - Cajal, S. R. Y. e L. Azoulay (1909). Histologie du système nerveux de l’homme et des vertébrés. Parigi, Maloine.
- 4.0 4.1 Kandel, et al Principles of Neuroscience. Quarta edizione. pp 591-624. Copyright 2000, da McGraw-Hill Co. Citare errore: Invalid
<ref>tag; name “R2” defined multiple times with different content - Kulesza, R. J., Jr. (2007). “Citoarchitettura del complesso olivario superiore umano: oliva superiore mediale e laterale”. Hear Res 225(1-2): 80-90.
- Tsuchitani, C. e J. C. Boudreau (1967). “Codifica della frequenza e dell’intensità dello stimolo da parte delle cellule del segmento S dell’oliva superiore del gatto”. J Acoust Soc Am 42(4): 794-805.
- 8.0 8.1 8.2 Citare errore: Invalid
<ref>tag; name “R10” defined multiple times with different content - Wu, S. H. and J. B. Kelly (1994). “Prove fisiologiche per l’inibizione omolaterale nell’oliva superiore laterale: risposte sinaptiche in fetta di cervello di topo”. Hear Res 73(1): 57-64.
- Brownell, W. E., P. B. Manis, et al. (1979). “Risposte inibitorie omolaterali nell’oliva laterale superiore del gatto”. Brain Res 177(1): 189-93.
- Darrow, K. N., S. F. Maison, et al. (2006). “Il feedback efferente cocleare bilancia la sensibilità interaurale”. Nat Neurosci 9(12): 1474-6.
- Warr, W. B. e J. J. Guinan, Jr. (1979). “Innervazione efferente dell’organo del corti: due sistemi separati”. Brain Res 173(1): 152-5.
- 13.0 13.1 Schofield, B. R. e N. B. Cant (1991). “Organizzazione del complesso olivario superiore nella cavia. I. Citoarchitettura, istochimica del citocromo ossidasi e morfologia dendritica”. J Comp Neurol 314 (4): 645-70. Citare errore: Invalid
<ref>tag; name “R7” defined multiple times with different content - 14.0 14.1 14.2 14.3 14.4 14.5 14.6 14.7 14.8 Illing, R. B., K. S. Kraus, et al. (2000). “Plasticità del complesso olivario superiore”. Microsc Res Tech 51(4): 364-81. Citare errore: Invalid
<ref>tag; nome “R5” definito più volte con contenuto diverso Citare errore: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; nome “R5” definito più volte con contenuto diverso - Warr, W. B. e J. E. Beck (1996). “Proiezioni multiple dal nucleo ventrale del corpo trapezoidale nel ratto”. Hear Res 93(1-2): 83-101.
- 16.00 16.01 16.02 16.03 16.04 16.05 16.06 16.07 16.08 16.09 16.10 Schofield, B. R. e N. B. Cant (1992). “Organizzazione del complesso olivario superiore nella cavia: II. Modelli di proiezione dai nuclei periolivari al collicolo inferiore”. J Comp Neurol 317 (4): 438-55. Cite error: Invalid
<ref>tag; nome “R6” definito più volte con contenuto diverso Cite error: Invalid<ref>tag; nome “R6” definito più volte con contenuto diverso Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content - Smith, P. H., P. X. Joris, et al. (1993). “Proiezioni di assoni di cellule sferiche a cespuglio fisiologicamente caratterizzate dal nucleo cocleare del gatto: evidenza di linee di ritardo verso l’oliva superiore mediale.” J Comp Neurol 331(2): 245-60.
- Wenthold, R. J., D. Huie, et al. (1987). “Immunoreattività della glicina localizzata nel nucleo cocleare e nel complesso olivario superiore”. Neuroscienze 22(3): 897-912.
- 19.0 19.1 Adams, J. C. (1983). “Citologia delle cellule periolivari e l’organizzazione delle loro proiezioni nel gatto”. J Comp Neurol 215(3): 275-89. Cite error: Invalid
<ref>tag; nome “R8” definito più volte con contenuto diverso - Friauf, E. e J. Ostwald (1988). “Proiezioni divergenti di neuroni del nucleo cocleare ventrale di ratto fisiologicamente caratterizzati come mostrato dall’iniezione intra-assonale di perossidasi di rafano”. Exp Brain Res 73(2): 263-84.
- Kulesza, R. J., Jr. e A. S. Berrebi (2000). “Il nucleo paraolivare superiore del ratto è un nucleo GABAergico”. J Assoc Res Otolaryngol 1(4): 255-69.
- Kulesza, R. J., Jr., G. A. Spirou, et al. (2003). “Proprietà di risposta fisiologica dei neuroni nel nucleo paraolivare superiore del ratto”. J Neurophysiol 89(4): 2299-312.
- WHITE JS, WARR WB. La doppia origine del fascio olivococcleare nel ratto albino. J. Comp. Neurol. 219:203-214, 1983.
- Guinan, J. J., Jr., S. S. Guinan, et al. (1972). “Singole unità uditive nel complesso olivario superiore. I. Risposte ai suoni e classificazioni basate su proprietà fisiologiche”. Int J Neurosci 4: 101-20.
- Tsuchitani, C. (1977). “Organizzazione funzionale dei gruppi cellulari laterali del complesso olivario superiore del gatto”. J Neurophysiol 40(2): 296-318.
- Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996). Origine embriologica per l’autismo: anomalie di sviluppo dei nuclei motori dei nervi cranici. J Comp Neurol 370 (2): 247-61.
- MedEd at Loyola Neuro/frames/nlBSs/nl27fr.htm
- Image at med.utah.edu (pick 9b. Pons)
Questo articolo era originariamente basato su una voce da un’edizione di pubblico dominio di Gray’s Anatomy. Come tale, alcune delle informazioni contenute possono essere obsolete. Si prega di modificare l’articolo se questo è il caso, e sentitevi liberi di rimuovere questo avviso quando non è più rilevante.
|
v-d-e
Cervello: rombencefalo (cervelletto) |
||||||
|---|---|---|---|---|---|---|
| Metencefalo/ pon |
|
Dorsale/
(tegmento) |
Ventrale/
(base) |
Raphe/reticular
|
||
| Mielencefalo/ medulla |
|
Dorsale
|
Ventrale
|
Raphe/reticolare
|
||
|
v-d-e
Sistema sensoriale: Sistemi uditivo e vestibolare (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Orecchio esterno |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Earlobe) – Canale auricolare – Muscoli auricolari Eardrum (Umbo, Pars flaccida) |
|||||||||||
| Orecchio medio |
|
|||||||||||
| Orecchio interno/ (labirinto membranoso, labirinto osseo) |
|
|||||||||||
| {| class=”navbox collapsible nowraplinks” style=”margin:auto;” | ||||||||||||
|
—
|
||||||||||||
|
||||||||||||
|}
Questa pagina usa contenuti di Wikipedia con licenza Creative Commons (visualizza autori).
Lascia un commento