Frontiers in Psychology
Il Settembre 28, 2021 da adminIl corpo calloso (CC) è la commissura primaria che collega i due emisferi cerebrali del cervello. Pertanto, mentre il termine commissurotomia non è errato per descrivere il cosiddetto “split-brain”, ed è strettamente corretto per le operazioni chirurgiche umane originali in cui tutte le commissure sono state tagliate (Gazzaniga et al, 1967), gli interventi successivi hanno reciso solo la CC, facendo così entrare in uso il termine “callosotomia” che rimane il termine più appropriato in quei casi (per esempio, Franz et al., 1996, 2000).
Da una prospettiva evolutiva, la CC è uno sviluppo recente osservato solo nei mammiferi placentari (Katz et al., 1983; Mihrshahi, 2006). Si pensa che la funzionalità fornita dal CC sia stata preceduta, almeno in parte, dalla commissura anteriore (AC), come osservato nei mammiferi non placentari come i marsupiali (Heath e Jones, 1971; Ashwell et al., 1996). Anche se alcune vie residue del CA sono ancora presenti nel cervello umano, la sua connettività e la struttura rimane poco studiata nella letteratura attuale e i risultati radi sono piuttosto incoerenti (Lasco et al., 2002; Patel et al., 2010; Choi et al., 2011). Per esempio, i risultati basati sulla risonanza magnetica (MRI) sono contrastanti sul fatto che il CA abbia dimensioni simili nelle femmine e nei maschi (Lasco et al., 2002; Patel et al., 2010). In uno studio relativamente recente utilizzando la tecnica in vivo di tensore di diffusione (DTI) che capitalizza la diffusione delle molecole d’acqua negli assoni, Patel et al. (2010) hanno studiato otto soggetti e isolato con successo le fibre del CA in cinque soggetti. Tra i risultati sono stati, in un soggetto una grande proporzione di fibre nell’arto posteriore del CA che viaggia bilateralmente nelle regioni parietali dove la maggior parte di loro termina, con un fascio più piccolo che termina nel giro precentrale, e un altro fascio che entra nel lobo temporale. Tali differenze individuali sono state trovate in tutti i partecipanti, ma la maggioranza dei soggetti ha mostrato proiezioni bilaterali alle cortecce orbitofrontali, parietali e temporali, di cui il CC non proietta chiaramente. Insieme, i risultati implicano che l’AC ha sia proiezioni primarie che vestigiali ad aree neocorticali dove il CC non proietta, come altri hanno anche suggerito (Di Virgilio et al., 1999; Patel et al., 2010).
A nostro avviso, l’AAA si basa fortemente su reti neocorticali e sulla connettività con strutture sottocorticali come i gangli della base e il talamo (tra gli altri non discussi specificamente nella nostra proposta) (Franz, 2012). Quindi, qualsiasi sviluppo evolutivo che coinvolge l’AC potrebbe anche avere qualche ruolo nel modello AAA. Tuttavia, abbiamo trovato poca letteratura che affronta specificamente questo possibile collegamento.
Il ruolo dell’AC è stato studiato in compiti che probabilmente richiedono forme di attenzione, come quelli che coinvolgono l’attenzione visiva. Per esempio, è noto che le popolazioni acallosali sono state studiate nel contesto di compiti che coinvolgono l’integrazione visiva (Gazzaniga et al., 1962, 1965; Sperry, 1968; Levy et al., 1972). Più recentemente, Corballis (1995) ha cercato di affrontare l’integrazione visiva e l’attenzione nel tentativo di spiegare le discrepanze precedenti nella ricerca acallosale. Corballis ha proposto un sistema di attenzione visiva dicotomico; una parte (che coinvolge la forma) è automatica e localizzata in ogni emisfero, e l’altra (che coinvolge il movimento e la posizione) è volontaria e sottocorticale. Il modello AAA può basarsi su questa osservazione e affrontare le discrepanze osservate in letteratura. Sulla base del nostro modello proposto, attraverso più livelli di attenzione ci sono percorsi diversi dal CC che potrebbero mediare il trasferimento di informazioni tra gli emisferi. Una possibilità coinvolge il circuito dei gangli della base che proponiamo sia potenzialmente critico nel mediare la condivisione dell’attenzione tra gli emisferi (Franz, 2012). Sezionando il CC il trasferimento diretto dell’attenzione attraverso le vie corticali è in gran parte abolito (a meno che, naturalmente, l’AC non svolga un ruolo di riserva). Tuttavia, l’integrazione delle informazioni visive basata sull’attenzione potrebbe ancora essere mediata da vie sottocorticali e/o dal CA. In questo modo il sistema di attenzione in due parti osservato da Corballis (1995) potrebbe essere presente a causa della mediazione dell’integrazione visiva basata sull’attenzione attraverso l’AC (e/o i gangli della base e i circuiti coinvolti: Franz, 2012).
Gli studi che coinvolgono la visione e l’attenzione non devono essere considerati completamente separati dal sistema che regola l’azione motoria. In base alle nostre scoperte (Franz, 2004; Franz e Packman, 2004), varie forme di attenzione (visiva e interno-non visiva) sono strettamente legate all’azione motoria, ed è logico che una possibile chiave per comprendere un potenziale ruolo di attenzione dell’AC risieda nel legame tra azione, visione e attenzione. In uno studio precedente, Franz (2004) ha fatto disegnare a partecipanti normali dei cerchi bimanuali sotto diverse manipolazioni del feedback visivo, o dell’attenzione interna a ciascuna mano. Le dimensioni dei cerchi disegnati da una mano che riceveva una forma di attenzione interna diventavano più grandi in modo simile agli effetti che si verificavano con la presenza di un feedback visivo di quella mano (rispetto a nessuna attenzione/ feedback visivo). Il nostro modello AAA implica che l’attenzione sia una componente chiave nella regolazione dell’output motorio (così come la selezione di quale azione motoria è pianificata), e per quanto riguarda le prove presenti, non possiamo escludere che l’AC svolga un ruolo in questa funzione. Tuttavia, prova diretta del ruolo del CA in qualsiasi compito cognitivo non è banale.
Testare se il CA prende parte nella funzione cognitiva è quasi impossibile nella ricerca neuropsicologica dato che i partecipanti con AC localizzato non sono facili da trovare (e non sappiamo di nessuno). Potrebbe quindi essere molto più intuitivo valutare le differenze neurologiche nel CA normale e correlare i dati neurologici con quelli comportamentali. Il metodo presentato tractografia (delucidando il CA) può essere applicato relativamente facilmente attraverso grandi campioni della popolazione normale, e idealmente, può essere combinato con le prestazioni (sulla base di compiti condotti durante le sessioni di test separati dalla raccolta DTI) di quegli stessi individui utilizzando compiti cognitivi appositamente progettato. Qui, dimostriamo che la DTI può essere condotta e può dare risultati utilizzabili su un piccolo campione di persone della popolazione neurologicamente normale. La proposta generale sarebbe quella di condurre metodi simili su una dimensione del campione più grande, e (come suggerito sopra), correlare i risultati (di una misura proxy della densità del CA, per esempio) con variabili su un compito cognitivo.
Nella Figura 1 mostriamo i risultati del monitoraggio DTI su dati ottenuti da 10 partecipanti femminili ottenuti da un set di dati DTI pubblico alla Johns Hopkins University (vedi Ringraziamenti per la fonte completa). L’età media era di 22 anni (SD = 2.5). Le immagini sono state elaborate utilizzando FSL Diffusion Toolbox (Smith et al., 2004; Woolrich et al., 2009; Jenkinson et al., 2012) e le procedure utilizzate in studi precedenti su altri tratti di fibre (FDT; Behrens et al., 2003a,b, 2007; Johansen-Berg et al., 2004). Brevemente, le immagini sono state prima concatenate in volumi 4d per ogni partecipante e crani spogliato utilizzando FSL Brain Extraction Tool (BET; Smith, 2002; Jenkinson et al., 2005). Le immagini estratte sono state corrette per il movimento minore della testa e le correnti parassite generate dai cambiamenti nella direzione del campo. I tensori di diffusione sono stati adattati per generare immagini che mostrano le direzioni di diffusione primaria e i livelli di FA in ogni voxel da utilizzare per le mappe di colore durante il mascheramento. Stima bayesiana dei parametri di diffusione ottenuti utilizzando tecniche di campionamento con fibre di attraversamento (BEDPOSTX; “X” che implica fibre di attraversamento) è stato condotto per generare una distribuzione di dimensione e direzione di diffusione contabilità per due fibre di attraversamento per condurre successivamente la trattografia attraverso voxel di diffusione.
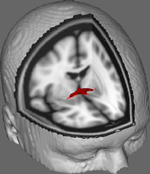
Figura 1. Media dei risultati di tracciamento in 10 partecipanti di controllo sovrapposti a un’immagine MRI ponderata T1 standard con voxel isotropi di 2 mm per riferimento anatomico.
I risultati medi di tracciamento in tutti i 10 partecipanti di controllo sono stati poi sovrapposti a un’immagine MRI ponderata T1 standard con voxel isotropi di 2 mm per riferimento anatomico. Il rapporto FA medio del gruppo era 0.3146 (SD = 0.0472). Questi risultati della trattografia rivelano, in modo importante, che la struttura del CA può essere chiarita, e quindi ha il potenziale per ulteriori studi che correlano i punteggi delle prestazioni su compiti comportamentali con i valori di FA utilizzando una dimensione del campione più grande.
In sintesi, al momento, esiste una scarsa letteratura, e della letteratura disponibile nessuno collega specificamente il CA ai processi relativi al modello AAA che è l’obiettivo del nostro articolo precedente (Franz, 2012). Utilizzando una procedura simile a quella qui proposta, è possibile testare campioni più grandi della popolazione normale, e correlare i valori FA dei risultati DTI (per esempio) con una specifica variabile dipendente ottenuta da un compito sviluppato ad hoc di attenzione all’azione (le cui procedure sono uno sviluppo in corso nel nostro laboratorio).
Conflict of Interest Statement
Gli autori dichiarano che la ricerca è stata condotta in assenza di relazioni commerciali o finanziarie che potrebbero essere interpretate come un potenziale conflitto di interessi.
Acknowledgments
Riconosciamo con gratitudine lo Human Brain Project e il National Research Resource Center (USA) e la John Hopkins University per aver permesso l’uso pubblico dei loro dati DTI. Siamo anche grati per il generoso sostegno del Marsden Fund della Royal Society of New Zealand (a Elizabeth A. Franz).
Corballis, M. (1995). Integrazione visiva nel cervello scisso. Neuropsicologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). L’allocazione dell’attenzione per l’apprendimento di azioni dirette all’obiettivo: un quadro di neuroscienze cognitive incentrato sui gangli della base. Front. Psychol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., e Jones, E. (1971). Percorsi interemisferici in assenza di un corpo calloso. Uno studio sperimentale delle connessioni commissurali nella falange marsupiale. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
Lascia un commento