Superior olivary nucleus
On november 12, 2021 by admin| Agy: | ||
|---|---|---|
 |
||
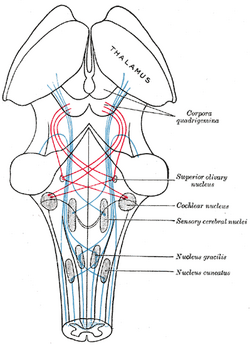
| Séma, amely a lemniscus rostjainak menetét mutatja; a mediális lemniscus kékkel, a laterális vörössel. (Jobbra középen a superior olivary nucleus van jelölve.) | ||
| ] | ||
| Latin | nucleus olivaris superior | |
| Gray | tárgy #187 787 | |
| részlete | ||
| Komponensek | ||
| Arteria | ||
| Véna | ||
| AgyInfo/UW | {{{AgyInfoTípus}}}-565 | |
| MeSH | ||
A felső olivari komplex (vagy SOC vagy superior olive) egy kis szürke anyagtömeg, amely a trapéztest laterális részének dorzális felszínén helyezkedik el. Emberben kicsi, de bizonyos állatokban jól fejlett, az alsó olivari maghoz hasonló felépítésű, és közvetlenül fölötte helyezkedik el. A trapéztest egyes rostjai e mag sejtjei körül arborizálódva végződnek, míg mások e sejtekből erednek.
Fiziológia
A felső olivari mag számos szerepet játszik a hallásban. A medialis superior olivárium (MSO) egy specializált mag, amely feltehetően a hangok fül közötti érkezési időkülönbség (interaurális időkülönbség vagy ITD) mérésére szolgál. Az ITD az alacsony frekvenciájú hangok azimutális irányának meghatározásában, azaz az azimutális síkban való lokalizálásukban – a balra vagy jobbra való irányuk mértékében – fontos támpont.
A laterális felső olajbogyó (LSO) feltehetően részt vesz a fülek közötti hangok szintkülönbségének (interaurális szintkülönbség vagy ILD) mérésében. Az ILD a második fő támpont a nagyfrekvenciás hangok azimutjának meghatározásában.
Kapcsolat a hallórendszerrel
A felső olivari komplex általában a ponsban található, de az emberben a rostrális medullától a középső agyvelőig terjed, és a ventrális akusztikus stria révén túlnyomórészt az anteroventralis cochleáris magból kap projekciókat, bár a posteroventralis mag is oda vetül a köztes akusztikus stria révén. Ezen a helyen történnek az első jelentős binaurális kölcsönhatások.
Elsődleges magok
A felső olivari komplexum három elsődleges magra, az MSO-ra, az LSO-ra és a trapéztest medialis magjára, valamint több kisebb periolivari magra oszlik. Ez a három mag a leginkább tanulmányozott, és ezért a legjobban megértett. Általában úgy tekintik, hogy ezek alkotják a felszálló azimutális lokalizációs pályát.
Medialis superior olive (MSO)
A medialis superior olive vélhetően segít lokalizálni a hang azimutját, vagyis azt a szöget balra vagy jobbra, ahol a hangforrás található. Az ember elsőre azt gondolhatja, hogy ez a mag magában foglalja a függőleges információt is, de ez nem így van; az MSO-ban feldolgozott információ a vízszintes síkra korlátozódik. A fusiform sejtek nem vetülnek ki semmire a pons szintjén, és csak az inferior colliculusnál lépnek működésbe. Csak horizontális adatok vannak jelen, de ezek két különböző fülforrásból származnak, ami segíti a hangok lokalizálását az azimutális tengelyen. A felsőbb oliva ezt úgy teszi, hogy az ugyanazt az ingert rögzítő két füljel közötti időkülönbségeket méri. A fej körüli utazás körülbelül 700 μs-t vesz igénybe, és feltételezzük, hogy a medialis superior oliva képes ezt érzékelni. Valójában megfigyelték, hogy az emberek akár 10 mikromásodpercig terjedő interaurális különbségeket is képesek érzékelni. A mag tonotopikusan szerveződik, de az azimutális receptív mező vetülete “valószínűleg egy komplex, nemlineáris térkép”.
A medialis superior oliva projekciói sűrűn végződnek az inferior colliculus ipsilateralis központi magjában. Ezen axonok többségét “kerek alakúnak” vagy R típusúnak tekintik. Ezek az R axonok többnyire glutamatergikusak, kerek szinaptikus vezikulákat tartalmaznak, és aszimmetrikus szinaptikus csomópontokat alkotnak.
- Ez a legnagyobb mag, és emberben körülbelül 15 500 neuront tartalmaz.
- Minden MSO alacsony frekvenciájú kétoldali bemenetet kap a jobb és bal AVCN-ból.
- A kimenet az ipsilaterális laterális lemniscusba és végül az inferior colliculusba megy.
- Az MSO jobban reagál a binaurális ingerekre.
- Fő funkciója a két fülhöz érkező hangok érkezési idejének különbségeinek detektálása, ami a lokalizációs folyamat (ITD) része.
Laterális superior oliva (LSO)
Ez az oliva hasonló funkciókkal rendelkezik, mint a medialis superior oliva, de az intenzitást használja a hangforrás lokalizálására. Ez az agytörzsnek az a része, amely a bal fül felől érkező hangosabb hangot a bal oldalon lévőnek jelöli. Az oldalsó oliva mindkét cochleáris magból, az ipsilaterális AVCN gömbölyded bokros sejtjeiből és a kontralaterális AVCN gömbölyded bokros sejtjeiből kap bemenetet az ipsilaterális MNTB-n keresztül. Az MNTB fő sejtjei glicinergek, gátlási forrást biztosítanak az LSO számára, és az ILD érzékenység alapját képezik. A mindkét cochleáris magból származó projekciók elsősorban magas frekvenciájúak, és ezeket a frekvenciákat később az LSO neuronok többsége képviseli (>2/3 2-3kHz felett macskában). További bemenetek származnak az ipsilaterális LNTB-ből (glicinerg, lásd alább), amelyek gátló információt szolgáltatnak az ipsilaterális cochleáris magból. Egy másik, valószínűleg gátló bemenet az ipsilaterális AVCN nem szférikus sejtjeiből származik. Ezek a sejtek vagy gömbölyded bokrosak, vagy többpólusúak (csillagszerűek). E két bemenet bármelyike lehet az alapja az ipsilaterális gátlásnak, amely az elsődleges gerjesztést kísérő válasz-térképeken látható, élesítve az egység frekvenciahangolását.
A LSO kétoldalt a colliculus inferior központi magjára (ICC) vetül. Az ipsilaterális projekciók elsősorban gátló (glicinerg), a kontralaterális pedig gerjesztő jellegűek. További projekciós célpontok közé tartozik a laterális lemniscus dorzális és ventrális magja (DNLL & VNLL). A DNLL-ből származó GABAerg projekciók képezik a GABA elsődleges forrását a halló agytörzsben, és kétoldalt az ICC-re és a kontralaterális DNLL-re vetülnek. Ezek a konvergáló gerjesztő és gátló kapcsolatok csökkenthetik az ILD-érzékenység szintfüggését az ICC-ben az LSO-hoz képest.
A további projekciók a Lateral Olivocochlear Bundle-t (LOC) alkotják, amely a cochleáris belső szőrsejteket innerválja. Ezek a projekciók feltehetően hosszú időállandóval rendelkeznek, és az egyes fülek által érzékelt hangszintek normalizálására szolgálnak, hogy segítsék a hangok lokalizációját. Jelentős faji különbségek léteznek: A LOC-projekciós neuronok rágcsálóknál az LSO-n belül helyezkednek el, ragadozóknál (pl. macska) pedig körülveszik az LSO-t.
Trapéztest medialis magja (MNTB)
- Az MNTB a SOC legkisebb része; az MNTB jelenléte az emberi agytörzsben kétséges.
- Minden neuron csak nagyfrekvenciás bemenetet kap a kontralaterális AVCN-ről tartott kagylón keresztül, és kimenete az ipsilaterális LSO-ra vetül.
- Kétféle választípus található: az AVCN-ben található orsósejtekhez hasonló “chopper típusú” és az AVCN-ben található bokorsejtekhez hasonló elsődleges típus.
Periolivari magok
A SOC hat-kilenc periolivari magból áll, az idézett kutatótól függően, amelyeket jellemzően az elsődleges magokhoz viszonyított elhelyezkedésük alapján neveznek el. Ezek a magok minden egyes elsődleges magot körülvesznek, és hozzájárulnak mind a felszálló, mind a leszálló hallórendszerhez. Ezek a magok képezik az olivocochlearis köteg forrását is, amely a csigát innerválja. Tengerimalacban a colliculi inferiorhoz vezető felszálló projekciók elsősorban ipsilaterálisak (>80%), a legnagyobb egyetlen forrás a SPON-ból származik. A ventrális magok (RPO, VMPO, AVPO, & VNTB) is szinte teljes egészében ipsilaterálisak, míg a többi mag kétoldali projekcióval rendelkezik.
| Név | Macska | Merimalac | Patkány | Eger | |||
|---|---|---|---|---|---|---|---|
| LSO | X | X | X | X | X | X | |
| MSO | X | X | X | X | |||
| MNTB | X | X | X | X | X | X | |
| LNTB | X | X | “LVPO” | X | |||
| ALPO | X | X | |||||
| PVPO | X | X | |||||
| PPO | X | X | “CPO” | ||||
| VLPO | X | ||||||
| DPO | X | X | X | ||||
| DLPO | X | X | |||||
| VTB | X | X | “MVPO” | X | |||
| AVPO | X | ||||||
| VMPO | X | X | |||||
| RPO | X | X | |||||
| SPN | “DMPO” | X | X | X |
Ventralis Nucleus of Trapezoid Body (VNTB)
- A VNTB az MNTB-től laterálisan elhelyezkedő kis mag, és az MSO-tól ventrálisan helyezkedik el.
- Ez a sejtek heterogén populációjából álló mag számos hallómagra vetül, és a mediális olivocochlearis köteget (MOC) alkotja, amely a cochleáris külső szőrsejteket innerválja . Ezek a sejtek elektromotilis rostokat tartalmaznak, és mechanikus erősítőként/csillapítóként működnek a cochleán belül.
- A mag mindkét IC-re vetül, kétoldali vetületű sejtek nincsenek.
A trapéztest laterális magja (LNTB)
- Az LSO ventralis részén található
- Az AVCN gömbölyded bokros sejtjei kétoldali kollaterálisokat, a globuláris bokros sejtek pedig ipsilaterális kollaterálisokat vetítenek az LNTB neuronokra.
- A sejtek immunreaktívak a glicinre, és retrográd módon jelölődnek tritiált glicin LSO-ba történő injektálását követően
- A sejtmag mindkét IC-re, kevés sejt kétoldali vetülettel, valamint az ipsilaterális LSO-ra vetül.
- A nagy multipoláris sejtek a cochleáris sejtmagra vetülnek, de az IC-re nem, mind macskában, mind tengerimalacban.
- A bemenet gyakran a tartott véggömbökön keresztül történik, ami nagyon gyors jelátvitelt eredményez.
Superior Periolivary Nucleus (SPON) (Dorsomedialis Periolivary Nucleus (DMPO))
- Az MNTB-től közvetlenül dorsalisan helyezkedik el
- Patkányban a SPON egy homogén GABAerg mag. Ezek a tonotopikusan szervezett neuronok gerjesztő bemenetet kapnak a kontralaterális ventrális cochleáris magban lévő octopus és multipoláris sejtektől, glicinerg (gátló) bemenetet az ipsilaterális MNTB-ből, egy ismeretlen GABAerg (gátló) bemenetet, és az ipsilaterális ICC-re vetülnek. A legtöbb neuron csak az inger eltolásakor reagál, 200 Hz-ig képes az AM ingerekre fázistolni, és ez képezheti az ICC időtartam-szelektivitásának alapját. Figyelemre méltó, hogy a SPON neuronok nem kapnak ereszkedő bemenetet az IC-ből, és nem vetül a cochleába vagy a cochleáris magba, mint sok periolivari mag.
- A tengerimalacban a kerek vagy ovális multipoláris sejtek mindkét IC-re vetülnek, sok sejt kétoldali vetülettel. A hosszúkásabb sejtek, amelyek a cochleáris magba vetülnek, nem vetülnek az ICC-be. Úgy tűnik, hogy két sejtpopuláció létezik, egy ipsilaterálisan és egy bilaterálisan projekciózó.
- A legtöbb információ a rágcsálók SPON-járól származott, a mag kiemelkedő mérete miatt ezeknél a fajoknál, nagyon kevés vizsgálatot végeztek macska DMPO-n, és egyik sem volt kiterjedt.
Dorsalis perioliváris mag (DPO)
- Az LSO-tól dorsalisan és medialisan helyezkedik el
- Egyaránt tartalmaz EE (mindkét fül által gerjesztett) és E0 (csak a kontralaterális fül által gerjesztett) egységeket.
- A neuronok tonotopikusan szervezettek és nagyfrekvenciájúak.
- Egyetlen maghoz tartozhat a DLPO-val együtt
- A mag mindkét IC-re vetül, kétoldalt nem vetülnek sejtek.
Dorsolaterális periolivari mag (DLPO)
- Az LSO-tól dorsalisan és laterálisan helyezkedik el
- Egyaránt tartalmaz EE (mindkét fül által gerjesztett) és E0 (csak a kontralaterális fül által gerjesztett) egységeket.
- A neuronok tonotopikusan szervezettek és alacsony frekvenciájúak.
- Egyetlen maghoz tartozhat a DPO-val együtt
- A mag mindkét IC-re vetül, kevés sejt vetül kétoldalt.
Ventrolaterális periolivari mag (VLPO)
- Az LSO ventrális hillusától ventralisan és azon belül helyezkedik el
- Egyaránt tartalmaz EI (kontralaterális fül által gerjesztett és ipsilaterális fül által gátolt) és E0 (csak a kontralaterális fül által gerjesztett) egységeket.
- A neuronok tonotopikusan szerveződnek, és magas frekvenciájúak.
Anterolaterális periolivari mag (ALPO)
- A mag mindkét IC-re vetül, kétoldalt nem vetülnek sejtek.
- A nagy multipoláris sejtek a cochleáris magba vetülnek, de az IC-be nem, mind a macskában, mind a tengerimalacban.
Ventromedialis periolivari mag (VMPO)
- Az MSO és az MNTB között helyezkedik el.
- Kétoldali projekciókat küld az ICC felé.
- A mag mindkét IC-be vetül, kétoldali vetületű sejtek nincsenek.
Rostrális periolivari mag (RPO) (Anterior periolivari mag (APO)).
- Az MSO rostrális pólusa és a VNLL között található
Caudális periolivari mag (CPO) (Posterior periolivari mag (PPO)).
- Az MSO caudalis pólusa és az arcmag (7N) között helyezkedik el
Posteroventralis periolivari mag (PVPO)
- A mag mindkét IC-re vetül, kétoldalt nem vetülnek sejtek.
Patofiziológia
Egy 21 éves autista, epilepsziás és mentális retardációban szenvedő nő boncolása során a superior olive közel teljes hiányát találták.
Lásd még
- Olivary body
- Kulesza RJ, Cytoarchitecture of the human superior olivary complex: Medial and lateral superior olive. Hearing Research 225(2007) 80-90
- 2.0 2.1 Oliver DL, et al. Axonal projections from the lateral and medial superior olive to the inferior colliculus of the cat: a study using electron microscopic autoradiography. J Comp Neurol. 1995 Sep 11;360(1):17-32 Cite error: Invalid
<ref>tag; name “R4” defined multiple times with different content - Cajal, S. R. Y. és L. Azoulay (1909). Histologie du système nerveux de l’homme et des vertébrés. Paris, Maloine.
- 4.0 4.1 Kandel, et al Principles of Neuroscience. Negyedik kiadás. pp 591-624. Copyright 2000, by McGraw-Hill Co. Cite error: Invalid
<ref>tag; name “R2” defined multiple times with different content - Kulesza, R. J., Jr. (2007). “Az emberi superior olivari komplexum citoarchitektúrája: mediális és laterális superior oliva:”. Hear Res 225(1-2): 80-90.
- Tsuchitani, C. és J. C. Boudreau (1967). “Az ingerek frekvenciájának és intenzitásának kódolása a macska superior olive S-szegmens sejtjei által”. J Acoust Soc Am 42(4): 794-805.
- 8.0 8.1 8.2 Cite error: Invalid
<ref>tag; name “R10” defined multiple times with different content - Wu, S. H. and J. B. Kelly (1994). “Az ipsilaterális gátlás fiziológiai bizonyítékai az oldalsó superior olivában: szinaptikus válaszok egér agyszeletben”. Hear Res 73(1): 57-64.
- Brownell, W. E., P. B. Manis, et al. (1979). “Ipsilaterális gátló válaszok a macska laterális superior olivában”. Brain Res 177(1): 189-93.
- Darrow, K. N., S. F. Maison, et al. (2006). “A cochleáris efferens visszacsatolás kiegyensúlyozza az interaurális érzékenységet”. Nat Neurosci 9(12): 1474-6.
- Warr, W. B. és J. J. Guinan, Jr. (1979). “A corti szerv efferens innervációja: két különálló rendszer”. Brain Res 173(1): 152-5.
- 13.0 13.1 Schofield, B. R. és N. B. Cant (1991). “A superior olivari komplex szerveződése tengerimalacban. I. Cytoarchitektúra, citokróm-oxidáz hisztokémia és dendritikus morfológia”. J Comp Neurol 314(4): 645-70. Cite error: Invalid
<ref>tag; name “R7” defined multiple times with different content - 14.0 14.1 14.2 14.3 14.4 14.5 14.6 14.7 14.8 Illing, R. B., K. S. Kraus, et al. (2000). “A felső olivari komplexum plaszticitása”. Microsc Res Tech 51(4): 364-81. Cite error: Invalid
<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content Cite error: Invalid<ref>tag; name “R5” defined multiple times with different content - Warr, W. B. and J. E. Beck (1996). “Multiple projections from the ventral nucleus of the trapezoid body in the rat”. Hear Res 93(1-2): 83-101.
- 16.00 16.01 16.02 16.03 16.04 16.05 16.06 16.07 16.08 16.09 16.10 Schofield, B. R. és N. B. Cant (1992). “A superior olivari komplex szerveződése tengerimalacban: II. A periolivari magokból az inferior colliculusba történő projekció mintázatai”. J Comp Neurol 317(4): 438-55. Cite error: Invalid
<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content Cite error: Invalid<ref>tag; name “R6” defined multiple times with different content - Smith, P. H., P. X. Joris, et al. (1993). “Fiziológiailag jellemzett gömbölyded bokorsejtek axonjainak projekciói a macska cochleáris magjából: bizonyíték késleltetési vonalakra a medialis superior oliva felé.” J Comp Neurol 331(2): 245-60.
- Wenthold, R. J., D. Huie, et al. (1987). “A cochleáris magban és a felső olivari komplexben lokalizált glicin immunreaktivitás”. Neuroscience 22(3): 897-912.
- 19.0 19.1 Adams, J. C. (1983). “A periolivari sejtek citológiája és projekcióik szerveződése a macskában”. J Comp Neurol 215(3): 275-89. Cite error: Invalid
<ref>tag; name “R8” defined multiple times with different content - Friauf, E. and J. Ostwald (1988). “A fiziológiailag jellemzett patkány ventrális cochleáris mag neuronjainak divergens projekciói, amint azt a torma-peroxidáz intra-axonális injekciója mutatja”. Exp Brain Res 73(2): 263-84.
- Kulesza, R. J., Jr. és A. S. Berrebi (2000). “A patkány superior paraolivari magja egy GABAerg mag”. J Assoc Res Otolaryngol 1(4): 255-69.
- Kulesza, R. J., Jr., G. A. Spirou, et al. (2003). “A patkány felső paraolivari magjának neuronjainak élettani válasz tulajdonságai”. J Neurophysiol 89(4): 2299-312.
- WHITE JS, WARR WB. Az olivocochlearis köteg kettős eredete az albínó patkányban. J. Comp. Neurol. 219:203-214, 1983.
- Guinan, J. J., Jr., S. S. Guinan, et al. (1972). “Egyedi hallási egységek a felsőbb olivari komplexben. I. Hangokra adott válaszok és fiziológiai tulajdonságokon alapuló osztályozások”. Int J Neurosci 4: 101-20.
- Tsuchitani, C. (1977). “A macska superior olivari komplexum laterális sejtcsoportjainak funkcionális szerveződése”. J Neurophysiol 40(2): 296-318.
- Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996). Az autizmus embriológiai eredete: az agyidegek motoros magjainak fejlődési rendellenességei. J Comp Neurol 370 (2): 247-61.
- MedEd at Loyola Neuro/frames/nlBSs/nl27fr.htm
- Image at med.utah.edu (pick 9b. Pons)
Ez a cikk eredetileg a Gray’s Anatomy public domain kiadásának bejegyzésén alapul. Ezért az itt szereplő információk egy része elavult lehet. Kérjük, szerkessze a cikket, ha ez a helyzet, és bátran távolítsa el ezt a bejegyzést, ha már nem releváns.
|
v-d-e
Agy: |
||||||
|---|---|---|---|---|---|---|
| Metencephalon/ pons |
|
Dorsalis/
(tegmentum) |
Ventralis/
(bázis) |
Raphe/retikuláris
|
||
| Myelencephalon/ medulla |
|
Dorsalis
|
Ventralis
|
Raphe/retikuláris
|
||
|
v-d-e
Érzékszervi rendszer: (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| külső fül |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Earlobe) – Hallójárat – Auricularis izmok Eardrum (Umbo, Pars flaccida) |
|||||||||||
| középfül |
|
|||||||||||
| Belsőfül/ (hártyás labirintus, csontos labirintus) |
|
|||||||||||
| {| class=”navbox collapsible nowraplinks” style=”margin: | ||||||||||||
|
—
|
||||||||||||
|
||||||||||||
|}
Ez az oldal a Wikipédia Creative Commons licencelt tartalmát használja (szerzők megtekintése).
Vélemény, hozzászólás?