” Mekkora egy E. coli sejt és mekkora a tömege?
On december 3, 2021 by adminMekkora egy E. coli sejt és mekkora a tömege?
Olvasó mód
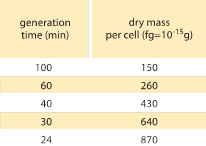
Táblázat 1: A baktérium tömege és az osztódási idő közötti összefüggés. Az egy sejtre jutó száraz tömeg a generációs (megduplázódási) idő függvényében van megadva. A tömeg a javaslat szerint nagyjából exponenciálisan növekszik a növekedési sebességgel, ahogyan azt eredetileg M. Schaechter et al. megfigyelte J. Gen. Microbiol., 19:592, 1958. A sejtek száraz tömegét 173 µg per egy mL OD460 egységre vonatkoztatott érték (BNID 106437) alapján számoltuk ki. A felhasznált törzs a B/r, a korai bakteriális fiziológiai vizsgálatokban általánosan használt törzs. Az értékek F. C. Neidhardt, “Escherichia coli and Salmonella: Cellular and Molecular Biology”, Vol. 1., Chapter 3, ASM Press, 1996.
A tipikus baktérium, például az E. coli mérete kényelmes standard vonalzónak szolgál a molekuláris és sejtbiológiában a hosszskálák jellemzésére. A fény- és elektronmikroszkópos mérések generációin alapuló “ökölszabály” az E. coli sejt méreteire vonatkozóan az, hogy az átmérője körülbelül ≈1µm, a hossza ≈2µm, a térfogata pedig ≈1µm3 (1 fL) (BNID 101788). Az alakot gömbhengerként, azaz félgömb alakú kupakkal ellátott hengerrel közelíthetjük meg. Az idézett átmérő és hosszúság ismeretében kiszámíthatjuk a térfogat finomabb becslését ≈1,3 µm3 (5π/12, hogy pontosak legyünk). Az ezen érték és a fent idézett ökölszabály szerinti érték közötti különbség mutatja, hogy milyen mértékű következetlenséggel élünk együtt kényelmesen az ökölszabályok használatakor. Az egyik legegyszerűbb út egy baktérium tömegének becsléséhez az, hogy kihasználjuk egy E. coli sejt ≈1 µm3 térfogatát, és feltételezzük, hogy sűrűsége megegyezik a vízével. Ez a naiv becslés egy másik standard értéket eredményez, nevezetesen azt, hogy egy olyan baktérium, mint az E. coli tömege ≈1 pg (pico=10-12). Mivel a legtöbb sejt körülbelül 2/3-ban vízből áll (BNID 100044, 105482), és a többi összetevő, például a fehérjék jellemző sűrűsége körülbelül a víz sűrűségének 1,3-szorosa (BNID 101502, 104272), a sejttérfogatról tömegre történő átváltás körülbelül 10%-os pontosságú.
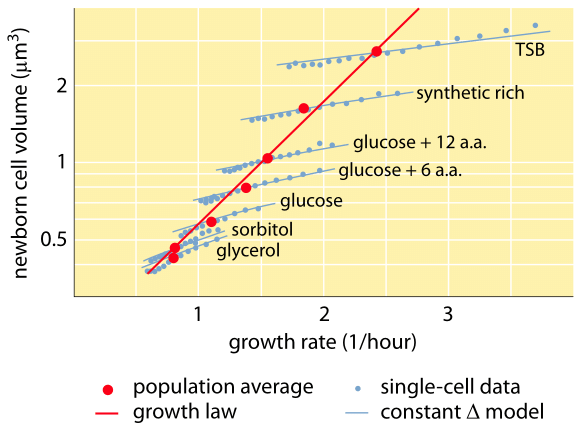
1. ábra: A sejttérfogat és a növekedési sebesség közötti összefüggés. Mikroszkópiával és mikrofluidikai eszközökkel a sejttérfogat egysejtes szinten mérhető különböző körülmények között, ami megerősíti, hogy az átlagos sejttérfogat exponenciálisan nő a növekedési sebességgel. Ezzel szemben a sejtek közötti eltérés egy adott feltétel esetén eltérő mértékben skálázódik. Az egysejtes viselkedés variációját a sejtméret-szabályozás modelljeinek tesztelésére használjuk. (Adaptálva S. Taheri-Araghi et al., Curr. Biol. 25:385, 2015.)
A bakteriális fiziológia egyik klasszikus eredménye hangsúlyozza, hogy a sejtek tulajdonságainak plaszticitása a sejttömeg növekedési sebességtől való függéséből ered. Egyszerűen megfogalmazva, a gyorsabb növekedési sebesség nagyobb sejtekhez társul. Ez a megfigyelés olyan fiziológiai változásokra utal, ahol a növekedési sebességet növelő közegek nagyobb sejteket is eredményeznek, amint azt az 1. ábra mutatja. Ez genetikailag is igaznak bizonyult, ahol a gyorsabb növekedési sebességet eredményező hosszú távú kísérleti evolúciós vizsgálatok nagyobb sejttérfogatot mutattak (BNID 110462). Az ilyen megfigyelések segítenek eloszlatni a “sejt” mítoszát – amikor az emberek, gyakran akaratlanul, egy sejtre vonatkozó méréseket használnak arra, hogy más sejttípusokra vagy ugyanarra a sejttípusra más körülmények között következtetéseket vonjanak le. Dennis és Bremer klasszikus tanulmányai rendszerezték ezeket a méréseket, és megállapították, hogy a száraz tömeg az 1. táblázatban látható módon változik a 100 percenként osztódó sejtek 148 fg-os átlagértékétől a 24 perces osztódási idővel rendelkező sejtek 865 fg-os értékéig, ami több mint ötszörös különbséget jelez a növekedési sebességtől függően. Hasonló tendenciát tapasztaltak más szervezeteknél is (pl. a bimbózó élesztő esetében, BNID 105103). Körülbelül 70%-os vízszint mellett ezek az értékek körülbelül 0,4 és 2,5 µm3 közötti tartománynak felelnek meg a térfogatot tekintve. Hogyan racionalizálhatjuk a nagyobb méreteket a gyorsabb ütemben növekvő sejtek esetében? Ez a kérdés a mai napig vita tárgya (Molenaar D. et al. MSB 5:323, 2009; Amir, A., Phys. Rev, Let., 112:208102, 2014). A magyarázatok változóak attól kezdve, hogy az erőforrás-elosztás módjának előnyére utalnak, egészen addig, hogy azt állítják, hogy ez valójában csak egy mellékhatása annak, hogy van egy körülbelül 60 perces beépített időszak attól kezdve, hogy a sejt úgy dönt, elég tömeget halmozott fel ahhoz, hogy megkezdje az osztódás előkészületeit, egészen addig, amíg befejezi a DNS-replikációt és az osztódás aktusát. Ez a nagyjából állandó “késleltetési” időszak ebben a gondolatmenetben a sejtek átlagos tömegének a növekedési sebességtől való exponenciális függéséhez vezet (Amir, A., Phys. Rev, Let.., 112:208102, 2014).
A sejttérfogat mérésének módszerei a Coulter-számláló használatától ((BNID 100004), amely egy kis nyílás ellenállásának változása alapján következtet a térfogatra, amikor egy sejt áthalad rajta, a közvetlenebb fluoreszcens mikroszkópiás mérésekig terjednek, amelyek különböző körülmények között mérik a sejtek hosszát és átmérőjét (1. ábra és BNID 106577, 111480). Meglepő módon a kalibrációs módszerek vagy a pontos törzsek és növekedési körülmények közötti különbségek miatt lehet, hogy a különböző laboratóriumok nem mindig közelítik meg ugyanazokat az értékeket. A sejttömeg mérésének eddig példa nélküli képességét a sejtek mikroszkopikus konzolon történő hatékony mérlegelésével érjük el. Amint azt a 2A. ábra szemlélteti, a folyadékáramlás segítségével a sejtet előre-hátra kényszerítik a kivájt konzolban. A mérés kihasználja azt a tényt, hogy a sejt tömege befolyásolja a konzol rezgési frekvenciáját. Ez a frekvencia fenomenális pontossággal mérhető, és femtogrammos pontossággal lehet belőle tömegekre következtetni. A folyadékáramlás irányának megváltoztatásával a sejtet percekig vagy még tovább is csapdába lehet ejteni, és a tömegfelhalmozódás mértéke egysejtes szinten folyamatosan mérhető. E technika első alkalmazása során kimutatták, hogy a nagyobb méretű egysejtűek gyorsabban halmozzák fel a tömeget, megvilágítva ezzel egy régóta fennálló kérdést: a sejtek növekedése lineáris az idővel, vagy inkább egy megközelítőleg exponenciális trenddel írható le? A különbségek parányiak lehetnek, de ezekkel a forradalmi képességekkel világosan látszott, hogy az utóbbi forgatókönyv jobban tükrözi a helyzetet több vizsgált sejttípus esetében, amint azt a 2B. ábra mutatja.
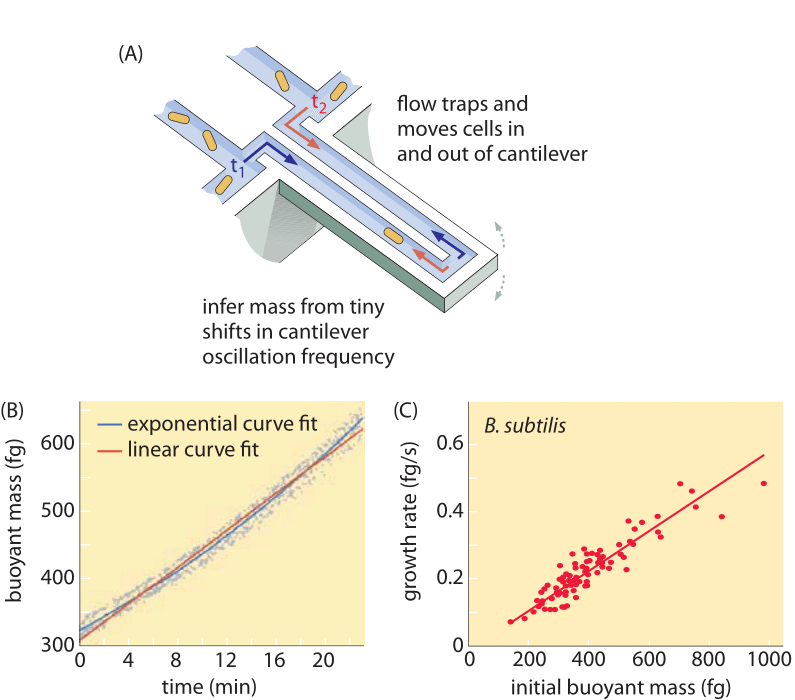
2. ábra: A felhajtó tömeg használata az egyes sejtek növekedésének mérésére. (A) Egy mikrométeres méretű konzol nagy frekvencián rezeg, és a rezgési frekvencia változásából meghatározható a sejtek tömege. (B) Az idő múlásával mérve ez egy egyedi sejtek tömegének felhalmozódási görbéjét eredményezi az ábrán látható módon. (C) Az ábrán B. subtilis sejtek láthatóak. A lineáris és az exponenciális növekedési modellek előrejelzéseinek összehasonlítása látható a legjobb illeszkedésként. A hasonlóság azt mutatja, hogy a két modell mennyire közel áll egymáshoz a sejtciklus során mindössze kétszeres növekedés tartományában. A sejtek száraz tömege körülbelül 4-szerese a felhajtó tömegnek. (Adapted from M. Godin et al., Nature Meth. 7:387, 2010.)





Vélemény, hozzászólás?