Hébi elmélet
On december 14, 2021 by admin Értékelés | Biopszichológia | Összehasonlító | Kognitív | Fejlődés | Nyelv | Egyéni különbségek | Személyiség | Filozófia | Szociális |
Módszerek | Statisztika | Klinikai | Oktatási | Oktatási | Ipari | Szakmai tételek | Világpszichológia |
Kognitív pszichológia:A Hebb-elmélet a szinaptikus plaszticitás alapvető mechanizmusát írja le, amelyben a szinaptikus hatékonyság növekedése a preszinaptikus sejt által a posztszinaptikus sejt ismételt és tartós ingerlésével jön létre. A Donald Hebb által 1949-ben bevezetett, Hebb-szabálynak, Hebb-posztulátumnak és sejtösszeszerelési elméletnek is nevezett elmélet a következőket mondja ki:
Tegyük fel, hogy egy reverzibilis aktivitás (vagy “nyom”) perzisztenciája vagy ismétlődése hajlamos olyan tartós sejtszintű változásokat előidézni, amelyek növelik a stabilitását…. Amikor az A sejt axonja elég közel van ahhoz, hogy egy B sejtet gerjesszen, és ismételten vagy tartósan részt vesz annak tüzelésében, akkor az egyik vagy mindkét sejtben valamilyen növekedési folyamat vagy anyagcsere-változás megy végbe, oly módon, hogy A mint a B-t tüzelő egyik sejt hatékonysága megnő.
Az elméletet gyakran úgy foglalják össze, hogy “a sejtek, amelyek együtt tüzelnek, össze vannak drótozva”, bár ez az idegrendszer túlzott leegyszerűsítése, amelyet nem szabad szó szerint venni, valamint nem tükrözi pontosan Hebb eredeti kijelentését a sejtek kapcsolódási erősségének változásáról. Az elméletet gyakran felidézik az asszociatív tanulás bizonyos típusainak magyarázatára, amelyekben a sejtek egyidejű aktiválása a szinaptikus erősség kifejezett növekedéséhez vezet. Az ilyen tanulást Hebb-tanulásnak nevezzük.
Hebb engrammok és a sejtösszekötő elmélet
A Hebb-elmélet azzal foglalkozik, hogy a neuronok hogyan kapcsolódhatnak össze, hogy engrammokká váljanak. Hebb elméletei a sejtösszefüggések formájáról és működéséről a következőkből érthetők meg:
“Az általános elképzelés régi, miszerint bármely két sejt vagy sejtrendszer, amelyek egyidejűleg ismételten aktívak, hajlamosak lesznek “társulni”, úgy, hogy az egyikben lévő aktivitás megkönnyíti a másikban lévő aktivitást”. (Hebb 1949, 70. o.) “Amikor az egyik sejt ismételten segíti a másik sejt tüzelését, az első sejt axonja szinaptikus csomókat fejleszt (vagy megnagyobbítja őket, ha már léteznek) a második sejt szomájával érintkezve”. (Hebb 1949, 63. o.)
Gordon Allport további gondolatokat vet fel a sejtösszeszerelési elmélettel és annak az engramok kialakulásában játszott szerepével kapcsolatban, az autoasszociáció fogalma mentén, amelyet a következőképpen ír le:
“Ha egy rendszer bemenetei miatt ugyanaz az aktivitási minta ismétlődik, az ezt a mintát alkotó aktív elemek halmaza egyre erősebben fog egymáshoz kapcsolódni. Azaz minden egyes elem hajlamos lesz bekapcsolni minden más elemet, és (negatív súlyokkal) kikapcsolni azokat az elemeket, amelyek nem képezik a mintázat részét. Másképpen fogalmazva, a minta egésze “önasszociálódik”. A megtanult (auto-asszociált) mintát nevezhetjük engrammnak”. (Hebb 1949, 44. o.)
A Hebb-elmélet volt az elsődleges alapja annak a hagyományos nézetnek, hogy holisztikus szinten elemezve az engrammok neuronhálózatok vagy neurális hálózatok.
Az Eric Kandel laboratóriumában végzett munka bizonyítékot szolgáltatott a Hebb-tanulási mechanizmusok szinapszisokban való részvételére a tengeri csigáknál (Aplysia californica).
A gerincesek központi idegrendszeri szinapszisainál a Hebb-féle szinapszismódosítási mechanizmusokkal kapcsolatos kísérleteket sokkal nehezebb ellenőrizni, mint a tengeri gerincteleneknél vizsgált viszonylag egyszerű perifériás idegrendszeri szinapszisokkal végzett kísérleteket. A gerinces neuronok közötti hosszú távú szinaptikus változásokkal (például a hosszú távú potenciálással) kapcsolatos munkák nagy része az agysejtek nem fiziológiai kísérleti ingerlésével történik. A gerincesek agyában vizsgált, fiziológiailag releváns szinapszismódosítási mechanizmusok némelyike azonban úgy tűnik, hogy a Hebb-folyamatok példája. Az egyik ilyen tanulmány olyan kísérletek eredményeit tekinti át, amelyek arra utalnak, hogy a szinaptikus erősségek hosszú távú változásait fiziológiailag releváns szinaptikus aktivitással lehet előidézni, amely mind Hebb-féle, mind nem Hebb-féle mechanizmusokon keresztül működik
Alapelvek
A mesterséges neuronok és mesterséges neurális hálózatok szempontjából a Hebb-elv a modellneuronok közötti súlyok módosításának meghatározási módjaként írható le. A két neuron közötti súly növekszik, ha a két neuron egyszerre aktiválódik – és csökken, ha külön-külön aktiválódnak. Azok a csomópontok, amelyek hajlamosak arra, hogy egyszerre mindkettő pozitív vagy mindkettő negatív legyen, erős pozitív súlyokkal rendelkeznek, míg azok, amelyek hajlamosak az ellentétes irányú aktivitásra, erős negatív súlyokkal rendelkeznek.
Ez az eredeti elv talán a súlyok kiválasztásának legegyszerűbb formája. Bár ez azt jelenti, hogy viszonylag könnyen be lehet kódolni egy számítógépes programba, és egy hálózat súlyainak frissítésére lehet használni, ugyanakkor a Hebbi-tanulás alkalmazásainak számát is korlátozza. Ma a Hebb-tanulás kifejezés általában a Hebb által javasolt eredeti elv valamilyen matematikai absztrakciójára utal. Ebben az értelemben a Hebb-tanulás során a tanulási csomópontok közötti súlyokat úgy módosítják, hogy az egyes súlyok jobban reprezentálják a csomópontok közötti kapcsolatot. Mint ilyen, számos tanulási módszer tekinthető némileg Hebb-féle természetűnek.
A következőkben a Hebb-tanulás képletes leírása következik: (megjegyezzük, hogy sok más leírás is lehetséges)
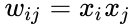
ahol 
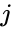
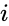
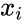
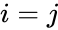
Egy másik képletszerű leírás a következő:
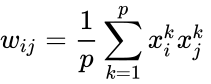
ahol 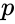

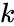
A Hebb-tanulás egy olyan változata, amely figyelembe veszi az olyan jelenségeket, mint a blokkolás és sok más neurális tanulási jelenség, Harry Klopf matematikai modellje. Klopf modellje nagyon sok biológiai jelenséget reprodukál, és ráadásul egyszerűen megvalósítható.
Általánosítás és stabilitás
A Hebb-szabályt gyakran általánosítják
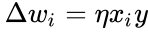
vagy a 
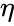
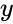
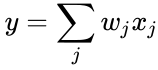
esetét, és az előző szakasz egyszerűsítése mind a tanulási rátát, mind a bemeneti súlyokat 1-nek veszi. A szabály ezen változata egyértelműen instabil, mivel bármely domináns jelű hálózatban a szinaptikus súlyok exponenciálisan nőnek vagy csökkennek. Megmutatható azonban, hogy bármilyen neuronmodell esetén a Hebb-szabály instabil. Ezért a neuronok hálózati modelljei általában más tanulási elméleteket alkalmaznak, mint például a BCM-elmélet, az Oja szabály vagy az általánosított Hebb-algoritmus.
Lásd még
- Anti-Hebbi tanulás
- BCM elmélet
- Coincidence Detection in Neurobiology
- Dale elve
- Generalizált Hebb-algoritmus
- Leabra
- Long.term potenciáció
- Memória
- Metaplaszticitás
- Neurális hálózatok
- Oja tanulási szabály
- Tetanikus stimuláció
- Spike-timing függő plaszticitás
- Synaptotrop hipotézis
- Shouval, Harel Az agy fizikája. A tanulás és a memória szinaptikus alapja: Egy elméleti megközelítés. The University of Texas Health Science Center at Houston. URL elérés: 2007-11-14.
További olvasmányok
- Hebb, D.O. (1949), The organization of behavior, New York: Wiley
- Hebb, D.O. (1961): Wiley
- Hebb, D.O. (1961). “Distinctive features of learning in the higher animal” J. F. Delafresnaye (szerk.) Brain Mechanisms and Learning, London: Oxford University Press.
- Hebb, D.O., and Penfield, W. (1940). Emberi viselkedés a homloklebenyek kiterjedt kétoldali eltávolítása után. Archives of Neurology and Psychiatry 44: 421-436.
- Allport, D.A. (1985). “Distributed memory, modular systems and dysphasia” Newman, S.K. and Epstein, R. (Eds.) Current Perspectives in Dysphasia, Edinburgh: Churchill Livingstone.
- Bishop, C.M. (1995). Neural Networks for Pattern Recognition (Neurális hálózatok a mintafelismeréshez), Oxford: Oxford University Press.
- Paulsen, O., Sejnowski, T. J. (2000). Természetes aktivitásminták és hosszú távú szinaptikus plaszticitás. Current opinion in neurobiology 10 (2): 172-179.
- Áttekintés
- Hebbian Learning tutorial (Part 1: Novelty Filtering, Part 2: PCA)
|
v-d-e
Neuroetológia |
||
|---|---|---|
| Koncepciók a neuroetológiában |
Feedforward – Coincidence detector -. Környezet – Ösztön – Jellemződetektor – Központi mintagenerátor (CPG) -NMDA receptor – Laterális gátlás – Rögzített cselekvési minta – Krogh-elv-Hebbi elmélet- Hanglokalizáció |
200px
|
| A neuroetológia története |
Theodore Holmes Bullock – Walter Heiligenberg -Niko Tinbergen- Konrad Lorenz- Eric Knudsen-Donald Griffin – Donald Kennedy – Karl von Frisch – Erich von Holst – Jörg-.Peter Ewert |
|
| Módszerek a neuroetológiában |
Whole Cell Patch Clamp |
|
| Modellrendszerek a neuroetológiában |
Animal Echolocation – Waggle… Dance- Electric Fish – Vision in toads – Neuroethology of Frog Audition – Infrared sensing in snakes |
|
This page uses Creative Commons Licensed content from Wikipedia (view authors).
Vélemény, hozzászólás?