Frontiers in Psychology
On szeptember 28, 2021 by adminA corpus callosum (CC) az agy két agyféltekéjét összekötő elsődleges kötőszövet. Ezért, bár a commissurotomia kifejezés nem helytelen az úgynevezett “osztott agy” leírására, és szigorúan helyes az eredeti emberi sebészeti műtétekre, amelyek során az összes commissurát elvágták (Gazzaniga et al., 1967), a későbbi műtétek során csak a CC-t vágták el, így került használatba a “callosotomia” kifejezés, amely ezekben az esetekben továbbra is a megfelelőbb kifejezés marad (pl. Franz et al., 1996, 2000).
Evolúciós szempontból a CC egy új keletű, csak a placentáris emlősöknél megfigyelhető fejlődés (Katz et al., 1983; Mihrshahi, 2006). A CC által biztosított funkcionalitást feltehetően – legalábbis részben – az elülső commissure (AC) előzte meg, ahogyan azt a nem placentáris emlősöknél, például az erszényesekben megfigyelték (Heath és Jones, 1971; Ashwell et al., 1996). Bár az AC néhány maradék útvonala még mindig jelen van az emberi agyban, konnektivitása és szerkezete a jelenlegi szakirodalomban továbbra is kevéssé tanulmányozott, és a ritka eredmények meglehetősen ellentmondásosak (Lasco et al., 2002; Patel et al., 2010; Choi et al., 2011). A mágneses rezonanciás képalkotáson (MRI) alapuló megállapítások például vegyesek abban a tekintetben, hogy az AC hasonló méretű-e a nőknél és a férfiaknál (Lasco et al., 2002; Patel et al., 2010). Egy viszonylag friss tanulmányban, amelyben a diffúziós tenzor képalkotás (DTI) in vivo technikáját használták, amely a vízmolekulák axonokban való diffúzióját használja fel, Patel és munkatársai (2010) nyolc személyt vizsgáltak, és öt személynél sikeresen izolálták az AC rostjait. A megállapítások között szerepelt, hogy az egyik alanyban az AC hátsó végtagjában lévő rostok nagy része kétoldalt a parietális régiókba haladt, ahol a legtöbbjük végződött, egy kisebb köteg a precentralis gyrusban végződött, és egy másik köteg pedig a temporális lebenybe lépett. Ilyen egyéni különbségeket találtunk minden résztvevőnél, mégis az alanyok többsége bilaterális projekciókat mutatott az orbitofrontális, a parietális és a temporális agykérgekbe, amelyek közül a CC nem vetül ki egyértelműen. Az eredmények együttesen azt sugallják, hogy az AAA-nak mind elsődleges, mind csökevényes projekciói vannak olyan neokortikális területekre, ahová a CC nem vetül, ahogy mások is javasolták (Di Virgilio et al., 1999; Patel et al., 2010).
A mi véleményünk szerint az AAA nagymértékben támaszkodik a neokortikális hálózatokra és a szubkortikális struktúrákkal, például a bazális ganglionokkal és a talamussal való kapcsolódásra (többek között a javaslatunkban külön nem tárgyaltak) (Franz, 2012). Így az AC-t érintő bármely evolúciós fejlődésnek is lehet némi szerepe az AAA-modellben. Azonban kevés irodalmat találtunk, amely kifejezetten ezzel a lehetséges kapcsolattal foglalkozik.
Az AC szerepét olyan feladatokban vizsgálták, amelyek valószínűleg a figyelem olyan formáit igénylik, mint például a vizuális figyelemmel kapcsolatos feladatok. Például jól ismert, hogy az acallosal populációkat vizuális integrációt igénylő feladatokkal összefüggésben tanulmányozták (Gazzaniga et al., 1962, 1965; Sperry, 1968; Levy et al., 1972). Nemrégiben Corballis (1995) a vizuális integrációval és a figyelemmel próbált foglalkozni, hogy megmagyarázza az acallosal-kutatás korábbi ellentmondásait. Corballis egy dichotóm vizuális figyelemrendszert javasolt; az egyik rész (amely a formát foglalja magában) automatikus és mindkét féltekére lokalizált, a másik (amely a mozgást és a helymeghatározást foglalja magában) pedig önkéntes és szubkortikális. Az AAA-modell erre a megfigyelésre építhet, és a szakirodalomban megfigyelt ellentmondásokat kezelheti. Az általunk javasolt modell alapján a figyelem több szintjén a CC-n kívül más útvonalak is léteznek, amelyek közvetíthetik a féltekék közötti információátadást. Az egyik lehetőség a bazális ganglionok áramköreit érinti, amelyek az általunk javasoltak szerint potenciálisan kritikusak a figyelem féltekék közötti megosztásának közvetítésében (Franz, 2012). A CC átvágásával a figyelem közvetlen átadása az agykérgi pályákon keresztül nagyrészt megszűnik (kivéve persze, ha az AC tartalék szerepet játszik). A vizuális információk figyelemalapú integrációja azonban továbbra is közvetíthető szubkortikális pályákon és/vagy az AC-n keresztül. Ily módon a Corballis (1995) által megfigyelt kétrészes figyelmi rendszer a vizuális integráció AC-n (és/vagy a bazális ganglionokon és az érintett áramkörökön keresztül történő figyelemalapú közvetítése miatt lehet jelen: Franz, 2012).
A látást és a figyelmet érintő tanulmányokat nem kell teljesen elkülönítve a motoros cselekvést szabályozó rendszertől vizsgálni. Saját eredményeink alapján (Franz, 2004; Franz és Packman, 2004) a figyelem különböző formái (vizuális és belső-nonvizuális) szorosan kapcsolódnak a motoros cselekvéshez, és logikus, hogy az AC lehetséges figyelmi szerepének megértéséhez az egyik lehetséges kulcs a cselekvés, a látás és a figyelem közötti kapcsolat. Egy korábbi vizsgálatában Franz (2004) normál résztvevőkkel rajzoltatott köröket kétkezi mozgással a vizuális visszacsatolás, illetve az egyes kezekre irányuló belső figyelem különböző manipulációi mellett. A belső figyelem egy formáját kapó kéz által rajzolt körök mérete nagyobb lett, hasonló módon, mint az adott kéz vizuális visszajelzésének jelenléte esetén fellépő hatások (a figyelem/vizuális visszajelzés hiányához képest). A mi AAA-modellünk a figyelmet a motoros kimenet szabályozásának kulcskomponenseként implikálja (valamint annak kiválasztását, hogy milyen motoros cselekvést tervezünk), és amennyiben a jelenlegi bizonyítékok alapján nem zárhatjuk ki, hogy az AC szerepet játszik ebben a funkcióban. Az AC bármely kognitív feladatban betöltött szerepének közvetlen tesztelése azonban nem triviális.
A neuropszichológiai kutatásokban szinte lehetetlen tesztelni, hogy az AC részt vesz-e a kognitív funkciókban, tekintettel arra, hogy lokalizált AC-vel rendelkező résztvevőket nem könnyű találni (és tudomásunk szerint nincs is). Ezért sokkal intuitívabb lehet a normális AC neurológiai különbségeinek felmérése és a neurológiai adatok viselkedési adatokkal való korrelálása. A bemutatott traktográfiás módszer (az AC megvilágítása) viszonylag könnyen alkalmazható a normál populáció nagy mintáin, és ideális esetben kombinálható ugyanezen egyének teljesítményével (a DTI-gyűjtésből különálló vizsgálati ülések során végzett feladatok alapján), célzott kognitív feladatokkal. A következőkben bemutatjuk, hogy a DTI elvégezhető és használható eredményeket adhat a neurológiailag normális populációból származó emberek kis mintáján. Az általános javaslat az lenne, hogy hasonló módszereket végezzünk nagyobb mintán, és (a fentiekben javasoltak szerint) korreláljuk az eredményeket (például az AC sűrűségének helyettesítő mértékét) egy kognitív feladat változóival.
Az 1. ábrán a DTI-követés eredményeit mutatjuk be 10 női résztvevőtől származó adatokon, amelyeket a Johns Hopkins Egyetem nyilvános DTI-adatkészletéből nyertünk (a teljes forrásért lásd a Köszönetnyilvánításban). Az átlagéletkor 22 év volt (SD = 2,5). A képeket az FSL Diffusion Toolbox (Smith et al., 2004; Woolrich et al., 2009; Jenkinson et al., 2012) segítségével dolgoztuk fel, és az eljárások a más rostpályákon végzett korábbi tanulmányokban (FDT; Behrens et al., 2003a,b, 2007; Johansen-Berg et al., 2004) alkalmazott eljárásokat alkalmazták. Röviden, a képeket először minden egyes résztvevő esetében 4d kötetekké konkatenálták, és a koponyákat az FSL Brain Extraction Tool (BET; Smith, 2002; Jenkinson et al., 2005) segítségével lecsupaszították. Az extrahált képeket korrigálták a fej kisebb mozgása és a mező irányának változásai által generált örvényáramok miatt. Diffúziós tenzorokat illesztettünk, hogy olyan képeket hozzunk létre, amelyek az egyes voxelekben az elsődleges diffúziós irányokat és FA-szinteket mutatják, amelyeket a maszkolás során színtérképekhez használhatunk. A keresztező rostokkal (BEDPOSTX; az “X” jelzi a keresztező rostokat) a mintavételi technikák segítségével kapott diffúziós paraméterek bayesi becslését végeztük el, hogy létrehozzuk a diffúzió méretének és irányának eloszlását, figyelembe véve a két keresztező rostot, hogy később a diffúziós voxeleken tractográfiát végezzünk.
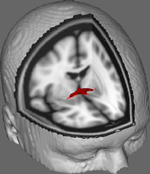
1. ábra. A 10 kontroll résztvevő átlagolt nyomkövetési eredményei egy standard T1-súlyozott MRI-képre helyezve, 2 mm-es izotróp voxelekkel az anatómiai referencia érdekében.
Az összes 10 kontroll résztvevő átlagolt nyomkövetési eredményeit ezután egy standard T1-súlyozott MRI-képre helyezve, 2 mm-es izotróp voxelekkel az anatómiai referencia érdekében. A csoport átlagos FA-aránya 0,3146 (SD = 0,0472) volt. Ezek a traktográfiás eredmények azt mutatják, ami fontos, hogy az AC szerkezete megvilágítható, és ezért potenciálisan további tanulmányok számára, amelyek a viselkedési feladatokon elért teljesítménypontszámokat korrelálják az FA-értékekkel, nagyobb mintanagyságot használva.
Összefoglalva, jelenleg kevés irodalom létezik, és a rendelkezésre álló irodalomból egyik sem kapcsolja kifejezetten az AC-t az AAA-modellel kapcsolatos folyamatokhoz, amely korábbi célcikkünk középpontjában áll (Franz, 2012). Az itt javasolthoz hasonló eljárás alkalmazásával lehetőség van a normál populáció nagyobb mintaszámú vizsgálatára, és a DTI-eredmények FA-értékeinek korrelációjára (például) egy konkrét függő változóval, amelyet a cselekvésre való figyelem célzottan kifejlesztett feladatából kapunk (amelynek eljárásai laboratóriumunkban folyamatban lévő fejlesztés).
Interdekütközésre vonatkozó nyilatkozat
A szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségként értelmezhetők.
Köszönet
Hálásan köszönjük a Human Brain Project és a National Research Resource Center (USA) és a John Hopkins Egyetem DTI-adatainak nyilvános használatának engedélyezését. Hálásak vagyunk továbbá az Új-Zélandi Királyi Társaság Marsden Alapjának nagylelkű támogatásáért (Elizabeth A. Franznak).
Corballis, M. (1995). Vizuális integráció az osztott agyban. Neuropsychologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). A figyelem elosztása a célzott cselekvések tanulására: egy kognitív idegtudományi keretrendszer a bazális ganglionokra összpontosítva. Front. Psychol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., and Jones, E. (1971). Interhemiszférikus pályák corpus callosum hiányában. A commissurális kapcsolatok kísérleti vizsgálata az erszényes phalangerben. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
Vélemény, hozzászólás?