A baktériumsejtek szerkezete és működése
On október 10, 2021 by adminA baktériumsejtek szerkezete és működése (5. oldal)
(Ez a fejezet 10 oldalas)
© Kenneth Todar, PhD
Cellafal
A baktériumok sejtfalai több okból is különleges figyelmet érdemelnek:
1. A fent leírtak szerint az életképességhez nélkülözhetetlen struktúra.
2. Olyan egyedi összetevőkből állnak, amelyek sehol máshol nem találhatók a természetben.
3. Az antibiotikumok támadásának egyik legfontosabb helyszínei.
4. Ligandumokat biztosítanak a tapadáshoz és receptorhelyeket a gyógyszerek vagy vírusok számára.
5. Az antibiotikumok és vírusok számára. Betegségtüneteket okoznak az állatokban.
6. Biztosítják a baktériumtörzsek közötti immunológiai megkülönböztetést és immunológiai variációt.
A legtöbb prokariótának merev sejtfala van.A sejtfal alapvető szerkezet, amely védi a sejtprotoplasztot a mechanikai károsodástól és az ozmotikus töréstől vagy lízistől.A prokarióták általában viszonylag híg környezetben élnek, így az oldott anyagok felhalmozódása a prokarióta sejt citoplazmájában jelentősen meghaladja a külső környezet teljes koncentrációját. Így a plazmamembrán belseje ellen ható ozmotikus nyomás 10-25 atm-nek felelhet meg. Mivel a membrán érzékeny, plasztikus szerkezet, nagy szakítószilárdságú, porózus, merev anyagból készült külső falnak kell visszatartania. Ilyen anyag a murein, a baktériumok sejtfalának mindenütt jelenlévő alkotóeleme.
A murein a peptidoglikán egy egyedülálló típusa, a diszacharidok (glikán) polimerje, amelyet rövid aminosavláncok (peptid) kötnek keresztbe. A peptidoglikánnak számos típusa létezik. Minden bakteriálispeptidoglikán N-acetil-muraminsavat tartalmaz, amely a murein meghatározó összetevője. Az Archaea sejtfalai fehérjékből, poliszacharidokból vagy peptidoglikánszerű molekulákból állhatnak, de soha nem tartalmaznak mureint. Ez a tulajdonság különbözteti meg a baktériumokat az archeáktól.
A Gram-pozitív baktériumokban (azokban, amelyek a Gram-festési eljárás során megtartják a lila kristályviola festéket) a sejtfal több réteg peptidoglikánból áll. A peptidoglikán rétegekre merőlegesen fut a teichonsavaknak nevezett molekulacsoport, amely csak a Gram-pozitív sejtfalra jellemző (14. ábra). 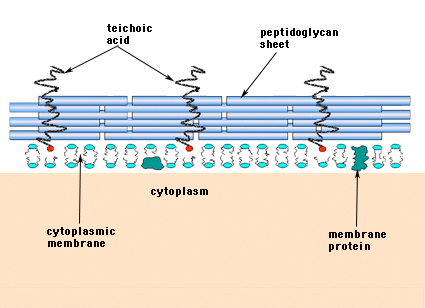
14. ábra. A Gram-pozitív baktérium sejtfalának szerkezete. A fal viszonylag vastag, és több peptidoglikán rétegből áll, amelyek között a peptidoglikán lapokra merőlegesen futó teichonsavak találhatók.
A Gram-negatív baktériumokban (amelyek nem tartják meg a kristályviolát) a sejtfal egyetlen réteg peptidoglikánból áll, amelyet egy külső membránnak nevezett hártyaszerkezet vesz körül.A Gram-negatív baktériumok külső membránja kivétel nélkül tartalmaz egy egyedi összetevőt, lipopoliszacharidot (LPS vagy endotoxin), amely az állatokra mérgező. A Gram-negatív baktériumoknál a külső membránt általában a sejtfal részének tekintik (15. ábra).

15. ábra. A Gram-negatív sejtfal felépítése. A fal viszonylagvékony, és sokkal kevesebb peptidoglikánt tartalmaz, mint a Gram-pozitív fal.a teichonsavak is hiányoznak. A Gram-negatív sejtfal azonban a peptidoglikánrétegen kívüli külső membránból áll. A külső membrán a peptidoglikán réteghez a lipoprotein molekulák egy sajátos csoportjával kapcsolódik.
A Gram-pozitív baktériumokban a sejtfal vastag (15-80nanométer),több peptidoglikán rétegből áll. A Gram-negatív baktériumokban a sejtfal viszonylag vékony (10 nanométer), és egyetlen peptidoglikán rétegből áll, amelyet külső membrán vesz körül.
Az E.coli peptidoglikán szerkezete és elrendezése reprezentatív az összes Enterobacteriaceae, valamint sok más Gram-negatív baktérium esetében. A glikán gerincet N-acetilglükozamin (G) és N-acetil-muraminsav (M) váltakozó molekulái alkotják, amelyeket béta-1,4-glikozid kötés köt össze. Az N-acetil-muraminsav (M) 3 szénatomja egy piruvátból származó laktil-éter csoporttal van szubsztituálva. A laktiléter a glikán gerincet egy peptid oldallánccal köti össze, amelyL-alanint (L-ala), D-glutamátot (D-glu), diaminopimélsavat (DAP) ésD-alanint (D-ala) tartalmaz. A MurNAc a baktériumok sejtfalában egyedülálló, csakúgy, mint aD-glu, a DAP és a D-ala. Az E. coli muraminsav alegysége az alábbi16. ábrán látható.

16. ábra. Az Escherichia coli peptidoglikánjának muraminsav alegységének szerkezete.ez a legtöbb Gram-negatív baktériumban megtalálható mureintípus. A glikángerinc két aminocukor, az N-acetilglükozamin (G) és az N-acetil-muraminsav (M) ismétlődő polimerje. Az N-acetil-muraminsavhoz egy trapeptid kapcsolódik, amely L-ala-D-glu-DAP-D-ala-ból áll. b. A muraminsav alegység rövidített szerkezete. c. A muraminsav alegység. A közeli tetrapeptid oldalláncok az egyik lánc DAP-ja és a másik lánc D-ala-ja közötti interpeptidkötéssel kapcsolódhatnak egymáshoz. d. A molekula polimer formája.
A mureinszálak a periplazmában kb. 10 muraminsav alegységből állnak össze. Ezután a szálak összekapcsolódnak, hogy egy folyamatos glikánmolekulát alkossanak, amely körülveszi a sejtet.Ahol a közelségük lehetővé teszi, a glikán gerincből kiálló tetrapeptidláncok a DAP szabad aminocsoportja és egy közeli D-ala szabad karboxicsoportja közötti interpeptidkötéssel keresztkötődhetnek.A peptidoglikán összeállását a plazmamembrán külső oldalán a periplazmatikus enzimek egy csoportja, a transzglikozilázok, transzpeptidázok és karboxipeptidázok közvetítik. A penicillin és a rokon béta-laktám antibiotikumok hatásmechanizmusa a transzpeptidáz és a karboxipeptidáz enzimek blokkolása a murein sejtfal összeszerelése során. Ezért mondják, hogy a béta-laktám antibiotikumok “blokkolják a sejtfalszintézist” a baktériumokban.
A peptidoglikánmolekula glikán gerincét a lizozim nevű enzim hasíthatja, amely jelen van az állati szérumban, a szövetekben és a váladékban, valamint a fagocita lizoszómában. A lizozim feladata a bakteriális sejtek lízise a bakteriális kórokozók elleni konstitutív védekezésként. Egyes Gram-pozitív baktériumok nagyon érzékenyek a lizozimre, és az enzim alacsony koncentrációban is igen aktív.A könnynedv (könny) 1:40 000 arányban hígítható, és megtartja a képességét, hogy bizonyos baktériumsejteket eltávolítson. A Gram-negatív baktériumok kevésbé érzékenyek a lizozim támadására, mivel peptidoglikánjukat a külső membrán védi. A lizozimális hasítás pontos helye a béta 1,4 kötés az N-acetil-muraminsav (M) és az N-acetilglükozamin (G) között, így a 16.(a) ábrán láthatóuraminsav alegység a lizozim bakteriális peptidoglikánon kifejtett hatásának eredménye.
A Gram-pozitív baktériumokban a peptidoglikánok között számos különböző peptidelrendezés létezik. A legjobban tanulmányozott a 17. ábrán bemutatott Staphylococcus aureusureinója. ADAP helyett (azE. coli-ban) a diaminsav, az L-lizin (L-lys), és az interpeptidkötés helyett (a Gram-negatívokban) egy aminosavakból álló interpeptidhíd, amely a lizin szabad aminocsoportját egy közeli tetrapeptid oldallánc D-ala szabad karboxicsoportjával köti össze. Ez az elrendezés nyilvánvalóan lehetővé teszi a közeli tetrapeptid-oldalláncok közötti gyakoribb keresztkötéseket. A S. aureusban az interpeptid-híd egy 5 glicinmolekulából álló peptid (úgynevezett pentaglicin-híd). A Gram-pozitív mureinben a béta-laktám antibiotikumok ugyanúgy gátolják az interpeptidhíd összeállását, mint a Gram-negatív mureinben az interpeptidkötést. A Gram-pozitív baktériumok érzékenyebbek a penicillintre, mint a Gram-negatív baktériumok, mivel a peptidoglikánt nem védi külső membrán, és ez egy nagyobb mennyiségben előforduló molekula. A Gram-pozitív baktériumokban a peptidoglikánok a tetrapeptid 3. pozíciójában a DAP vagy az L-lys helyén lévő aminosav, valamint az interpeptidhíd pontos összetételében változhatnak. A Gram-pozitív baktériumokban legalább nyolc különböző típusúpeptidoglikán létezik.
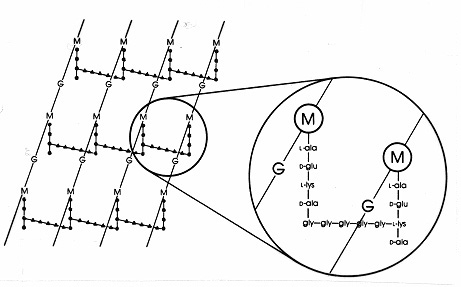
17. ábra. A Staphylococcus aureus peptidoglikán lapjának sematikus ábrája. G = N-acetil-glükozamin;M = N-acetil-muraminsav; L-ala = L-alanin; D-ala = D-alanin; D-glu= D-glutaminsav; L-lys = L-lizin. Ez a murein egyik típusa aGram-pozitív baktériumokban található. Az E. coli peptidoglikánhoz képest (7. ábra) a tetrapeptidben a DAP (diaminopimélsav) helyett L-lys található.Az L-lys szabad aminocsoportja egy glicin-pentapeptiddel (gly-gly-gly-gly-gly-gly-gly-) van helyettesítve, amely aztán egy interpeptidhíddá válik, amely egy szomszédos tetrapeptid oldalláncban lévő D-ala karboxicsoportjával alkot kapcsolatot. A Gram-pozitív peptidoglikánok fajonként különböznek,főként a tetrapeptidlánc harmadik pozíciójában lévő aminosavak és az interpeptid-híd aminosav-összetétele tekintetében.
A Gram-negatív baktériumok sejtfalában egyetlen murein-monomolekuláris réteg lehet, míg a Gram-pozitív baktériumok sejtfalában több peptidoglikán réteg vagy “burkolat” található. A Gram-pozitív baktériumok peptidoglikán rétegeihez szorosan kapcsolódik a teichonsavaknak nevezett molekulacsoport. A teichonsavak foszfátokkal és néhány aminosavval és cukorral helyettesített poliglicerin vagy poliribitol lineáris polimerjei. A teichonsav-polimerek esetenként a plazmamembránhoz rögzülnek (lipoteichonsavnak, LTA-nak nevezik), látszólag a peptidoglikán rétegekre merőlegesen kifelé irányítva. A teichoesav funkciói nem ismertek. Agram-pozitív baktériumok életképességéhez a természetben nélkülözhetetlenek. Az egyik elképzelés szerint egyenletesen orientált negatív töltésekből álló csatornát biztosítanak a pozitív töltésű anyagok átvezetéséhez a bonyolult peptidoglikán-hálózaton. Egy másik elmélet szerint a teichonsavak valamilyen módon részt vesznek a muraminsavegységek szabályozásában és összeszerelésében a plazmamembrán külső oldalán. Vannak olyan esetek, különösen a streptococcusokban, amikor a teichonsavakat a baktériumoknak a szöveti felületekhez való tapadásában vonták felelősségre.
fejezet folytatása
előző oldal
Az előző oldal
Vélemény, hozzászólás?