Frontiers in Psychology
On Setembro 28, 2021 by adminO corpus callosum (CC) é a comissura primária que liga os dois hemisférios cerebrais do cérebro. Portanto, enquanto o termo comissurotomia não é incorreto para descrever a chamada “split-brain”, e é estritamente correto para as operações cirúrgicas humanas originais nas quais todas as comissuras foram cortadas (Gazzaniga et al, 1967), as cirurgias posteriores cortaram apenas o CC, trazendo assim em uso o termo “calosotomia”, que permanece como o termo mais apropriado nesses casos (por exemplo, Franz et al., 1996, 2000).
De uma perspectiva evolutiva, o CC é um desenvolvimento recente observado apenas em mamíferos placentários (Katz et al., 1983; Mihrshahi, 2006). Pensa-se que a funcionalidade proporcionada pelo CC é precedida, pelo menos em parte, pela comissura anterior (AC), conforme observado em mamíferos não placentários como os marsupiais (Heath e Jones, 1971; Ashwell et al., 1996). Embora algumas vias residuais da CA ainda estejam presentes no cérebro humano, sua conectividade e estrutura permanece subestudada na literatura atual e os achados esparsos são bastante inconsistentes (Lasco et al., 2002; Patel et al., 2010; Choi et al., 2011). Por exemplo, os achados baseados na ressonância magnética (RM) são mistos quanto a se o CA é de tamanho semelhante em mulheres e homens (Lasco et al., 2002; Patel et al., 2010). Em um estudo relativamente recente utilizando a técnica in-vivo de difusão por imagem tensorial (DTI) que capitaliza a difusão de moléculas de água em axônios, Patel et al. (2010) estudaram oito sujeitos e isolaram com sucesso fibras do CA em cinco sujeitos. Entre os achados, em um sujeito, uma grande proporção de fibras no membro posterior do CA viajou bilateralmente para regiões parietais onde a maioria delas terminou, com um feixe menor terminando no giro pré-central, e ainda outro feixe entrando no lobo temporal. Tais diferenças individuais foram encontradas em todos os participantes, mas a maioria dos sujeitos mostrou projeções bilaterais para os cortices orbitofrontal, parietal e temporal, dos quais o CC não se projeta claramente. Em conjunto, as conclusões implicam que o CC tem projecções tanto primárias como vestigiais para áreas neocorticais onde o CC não projecta, como outros também sugeriram (Di Virgilio et al., 1999; Patel et al., 2010).
Na nossa opinião, o AAA assenta fortemente em redes neocorticais e conectividade com estruturas subcorticais como os gânglios basais e o tálamo (entre outros não discutidos especificamente na nossa proposta) (Franz, 2012). Assim, quaisquer desenvolvimentos evolutivos envolvendo a CA podem também ter algum papel no modelo AAA. Entretanto, encontramos pouca literatura abordando especificamente esta possível ligação.
O papel do CA tem sido estudado em tarefas que provavelmente requerem formas de atenção, tais como as que envolvem atenção visual. Por exemplo, é sabido que as populações acallosal têm sido estudadas no contexto de tarefas que envolvem integração visual (Gazzaniga et al., 1962, 1965; Sperry, 1968; Levy et al., 1972). Mais recentemente, Corballis (1995) procurou abordar a integração visual e a atenção num esforço para explicar as discrepâncias anteriores na pesquisa acallosal. Corballis propôs um sistema dicotômico de atenção visual; uma parte (envolvendo forma) é automática e localizada para cada hemisfério, e a outra (envolvendo movimento e localização) é voluntária e subcortical. O modelo AAA pode se basear nesta observação e abordar as discrepâncias observadas na literatura. Com base no nosso modelo proposto, através de múltiplos níveis de atenção existem outros caminhos para além do CC que podem mediar a transferência de informação entre hemisférios. Uma possibilidade envolve o circuito dos gânglios basais que propomos é potencialmente crítica na mediação da partilha da atenção entre hemisférios (Franz, 2012). Ao transitar o CC, a transferência directa de atenção através de vias corticais é largamente abolida (a menos, claro, que o CC desempenhe um papel de apoio). No entanto, a integração da informação visual baseada na atenção ainda pode ser mediada através de vias subcorticais e/ou da CA. Desta forma, o sistema de atenção em duas partes observado por Corballis (1995) poderia estar presente devido à mediação baseada na atenção da integração visual via CA (e/ou os gânglios basais e circuitos envolvidos: Franz, 2012).
Estudos envolvendo visão e atenção não precisam ser considerados inteiramente separados do sistema que regula a ação motora. Com base em nossas próprias descobertas (Franz, 2004; Franz e Packman, 2004), várias formas de atenção (visual e não visual) estão intimamente ligadas à ação motora, e é lógico que uma possível chave para entender um potencial papel de atenção da CA reside na ligação entre ação, visão e atenção. Em um estudo anterior, Franz (2004) tinha participantes normais desenhando círculos bimanualmente sob diferentes manipulações de feedback visual, ou atenção interna para cada mão. O tamanho dos círculos desenhados por uma mão recebendo uma forma de atenção interna tornou-se maior de forma similar aos efeitos que ocorreram com a presença de feedback visual dessa mão (em relação à ausência de feedback visual). Nosso modelo AAA implica a atenção como sendo um componente chave na regulação da saída motora (assim como a seleção do que a ação motora é planejada), e na medida em que a evidência presente, não podemos descartar o CA como desempenhando um papel nessa função. Entretanto, o teste direto do papel do CA em qualquer tarefa cognitiva não é trivial.
Testar se o CA participa da função cognitiva é quase impossível na pesquisa neuropsicológica, uma vez que participantes com CA localizada não são fáceis de encontrar (e nós não sabemos de nenhuma). Portanto, pode ser muito mais intuitivo avaliar diferenças neurológicas no CA normal e correlacionar dados neurológicos com dados comportamentais. O método de tractografia apresentado (elucidando o CA) pode ser aplicado com relativa facilidade em grandes amostras da população normal e, idealmente, pode ser combinado com o desempenho (com base em tarefas realizadas durante sessões de teste separadas da coleta do DTI) desses mesmos indivíduos usando tarefas cognitivas propositadamente projetadas. Aqui, demonstramos que o DTI pode ser conduzido e pode produzir resultados úteis em uma pequena amostra de pessoas da população neurologicamente normal. A proposta geral seria conduzir métodos similares em uma amostra maior, e (como sugerido acima), correlacionar os resultados (de uma medida proxy da densidade do AC, por exemplo) com variáveis sobre uma tarefa cognitiva.
Na Figura 1 mostramos resultados do rastreamento do DTI em dados obtidos de 10 participantes do sexo feminino, obtidos de um conjunto de dados públicos do DTI na Universidade Johns Hopkins (ver Agradecimentos para fonte completa). A média de idade foi de 22 anos (DP = 2,5). As imagens foram processadas usando a FSL Diffusion Toolbox (Smith et al., 2004; Woolrich et al., 2009; Jenkinson et al., 2012) e os procedimentos empregados em estudos anteriores sobre outros trajetos de fibras (FDT; Behrens et al., 2003a,b, 2007; Johansen-Berg et al., 2004). Resumidamente, as imagens foram concatenadas em volumes 4d para cada participante e os crânios foram removidos usando a Ferramenta de Extração Cerebral da FSL (BET; Smith, 2002; Jenkinson et al., 2005). As imagens extraídas foram corrigidas para movimentos menores da cabeça e correntes de redemoinho geradas por mudanças na direção do campo. Os tensores de difusão foram ajustados para gerar imagens que mostram as direções de difusão primária e os níveis de FA em cada voxel a serem usados para mapas coloridos durante o mascaramento. A estimativa Bayesiana dos parâmetros de difusão obtidos utilizando técnicas de amostragem com fibras de cruzamento (BEDPOSTX; “X” implicando fibras de cruzamento) foi conduzida para gerar uma distribuição de tamanho e direção de difusão contabilizando duas fibras de cruzamento para posteriormente conduzir a tractografia através de voxels de difusão.
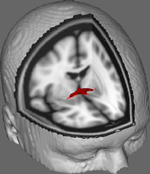
Figura 1. Resultados médios de rastreamento em 10 participantes de controle sobrepostos em uma imagem de RM padrão T1 com voxels isotrópicos de 2 mm para referência anatômica.
Resultados médios de rastreamento em todos os 10 participantes de controle foram então sobrepostos em uma imagem de RM padrão T1 com voxels isotrópicos de 2 mm para referência anatômica. A média do grupo FA foi de 0,3146 (DP = 0,0472). Estes resultados da tractografia revelam, de forma importante, que a estrutura do CA pode ser elucidada e, portanto, tem potencial para estudos adicionais correlacionando escores de desempenho em tarefas comportamentais com valores de FA usando uma amostra maior.
Em resumo, atualmente, existe uma literatura escassa, e da literatura disponível nenhuma liga especificamente o CA a processos relacionados ao modelo AAA que é o foco do nosso artigo alvo anterior (Franz, 2012). Usando um procedimento semelhante ao aqui proposto, é possível testar amostras maiores da população normal e correlacionar os valores de FA dos resultados do DTI (por exemplo) com uma variável dependente específica obtida a partir de uma tarefa de atenção à ação (procedimentos dos quais são um desenvolvimento contínuo em nosso laboratório).
Conflict of Interest Statement
Os autores declaram que a pesquisa foi conduzida na ausência de qualquer relação comercial ou financeira que pudesse ser interpretada como um potencial conflito de interesses.
Confirmações
Acreditamos com gratidão o Human Brain Project and National Research Resource Center (USA) e a John Hopkins University por permitirem o uso público de seus dados do DTI. Somos também gratos pelo generoso apoio do Fundo Marsden da Royal Society of New Zealand (para Elizabeth A. Franz).
Corballis, M. (1995). Integração visual no cérebro dividido. Neuropsicologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). A alocação de atenção à aprendizagem de acções orientadas por objectivos: um quadro neurocientífico cognitivo centrado nos gânglios basais. A frente. Psicol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., and Jones, E. (1971). Caminhos interhemisféricos na ausência de um corpus callosum. Um estudo experimental das conexões comissurais no falange marsupial. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
Deixe uma resposta