Structure et fonction des cellules bactériennes
On octobre 10, 2021 by adminStructure et fonction des cellules bactériennes (page 5)
(Ce chapitre a 10 pages)
© Kenneth Todar, PhD
Paroi cellulaire
Les parois cellulaires des bactéries méritent une attention particulière pour plusieurs raisons :
1. Elles sont une structure essentielle à la viabilité, comme décrit ci-dessus.
2. Elles sont composées de composants uniques que l’on ne trouve nulle part ailleurs dans la nature.
3. Elles sont l’un des sites les plus importants pour l’attaque par les antibiotiques.
4. Elles fournissent des ligands pour l’adhérence et des sites récepteurs pour les médicaments ou les virus.
5. Ils provoquent des symptômes de maladie chez les animaux.
6. Ils permettent une distinction immunologique et une variation immunologique entre les souches de bactéries.
La plupart des procaryotes ont une paroi cellulaire rigide.
La paroi cellulaire est une structure essentielle qui protège le protoplaste cellulaire des dommages mécaniques et de la rupture ou de la lyse osmotique.Les procaryotes vivent généralement dans des environnements relativement dilués, de sorte que l’accumulation de solutés à l’intérieur du cytoplasme de la cellule procaryote dépasse largement la concentration totale du milieu extérieur. Ainsi, la pression osmotique contre l’intérieur de la membrane plasmique peut être l’équivalent de 10-25atm. Comme la membrane est une structure plastique délicate, elle doit être retenue par une paroi extérieure faite d’un matériau poreux et rigide ayant une forte résistance à la traction. Un tel matériau est la mureine, le composant omniprésent des parois cellulaires bactériennes.
La mureine est un type unique de peptidoglycane, un polymère de disaccharides (glycan) réticulé par de courtes chaînes d’acides aminés (peptide). Il existe de nombreux types de peptidoglycane. Tous les peptidoglycanes bactériens contiennent de l’acide N-acétylmuramique, qui est le composant défini de la muréine. Les parois cellulaires des Archaea peuvent être composées de protéines, de polysaccharides ou de molécules de type peptidoglycane, mais elles ne contiennent jamais de muréine. Cette caractéristique distingue les bactéries des archées.
Dans les bactéries à Gram positif (celles qui retiennent le colorant violet de cristal violet lorsqu’elles sont soumises à la procédure de coloration de Gram), la paroi cellulaire est constituée de plusieurs couches de peptidoglycane. Les couches de peptidoglycane sont perpendiculaires à un groupe de molécules appelées acides teichoïques, qui sont propres à la paroi cellulaire des bactéries à Gram positif (figure 14). 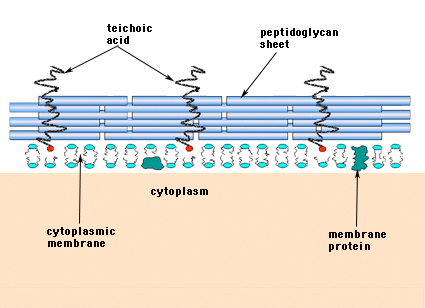
Figure 14. Structure de la paroi cellulaire des bactéries Gram-positives. La paroi est relativement épaisse et consiste en de nombreuses couches de peptidoglycane intercalées d’acides teichoïques qui s’étendent perpendiculairement aux feuillets de peptidoglycane.
Dans les bactéries Gram-négatives (qui ne retiennent pas le cristal violet), la paroi cellulaire est composée d’une seule couche de peptidoglycane entourée d’une structure membranaire appelée membrane externe.
La membrane externe des bactéries Gram-négatives contient invariablement un composant unique, le lipopolysaccharide (LPS ou endotoxine),qui est toxique pour les animaux. Chez les bactéries Gram-négatives, la membrane externe est généralement considérée comme faisant partie de la paroi cellulaire (figure 15).

Figure15. Structure de la paroi cellulaire des Gram-négatifs. La paroi est relativement mince et contient beaucoup moins de peptidoglycane que la paroi à Gram positif.De plus, les acides teichoïques sont absents. Cependant, la paroi des cellules Gram négatives est constituée d’une membrane externe qui se trouve à l’extérieur de la couche de peptidoglycane. La membrane externe est attachée à la feuille de peptidoglycane par un groupe unique de molécules lipoprotéiques.
Dans les bactéries Gram positives, la paroi cellulaire est épaisse (15-80nanomètres), constituée de plusieurs couches de peptidoglycane. Chez les bactéries à Gram négatif, la paroi cellulaire est relativement mince (10 nanomètres) et se compose d’une seule couche de peptidoglycane entourée d’une membrane externe.
La structure et l’agencement des peptidoglycanes chez E.coli sont représentatifs de tous les Enterobacteriaceae, ainsi que de nombreuses autres bactéries Gram-négatives. Le squelette du glycan est constitué de molécules alternées de N-acétylglucosamine (G) et d’acide N-acétylmuramique (M) reliées par une liaison bêta 1,4-glycoside. L’atome de carbone 3 de l’acide N-acétylmuramique (M) est substitué par un groupe éther lactylique dérivé du pyruvate. L’éther lactylique relie le squelette du glycan à une chaîne latérale peptidique qui contient de la L-alanine (L-ala), du D-glutamate (D-glu), de l’acide diaminopimélique (DAP) et de la D-alanine (D-ala). Le MurNAc est unique aux parois cellulaires bactériennes, tout comme le D-glu, le DAP et le D-ala. La sous-unité de l’acide muramique de E. coli est représentée dans laFigure16 ci-dessous.

Figure 16. La structure de la sous-unité de l’acide muramique du peptidoglycane d’Escherichia coli.C’est le type de muréine que l’on trouve dans la plupart des bactéries Gram-négatives. Le squelette du glycan est un polymère répété de deux sucres aminés, la N-acétylglucosamine (G) et l’acide N-acétylmuramique (M). L’acide N-acétylmuramique est rattaché à un tétrapeptide constitué de L-ala-D-glu-DAP-D-ala. b. Structure abrégée de la sous-unité de l’acide muramique. c. Les chaînes latérales tétrapeptidiques voisines peuvent être liées les unes aux autres par une liaison interpeptidique entre DAP sur une chaîne et D-ala sur l’autre. d. La forme polymère de la molécule.
Les brins de mureine sont assemblés dans le périplasme à partir d’environ 10 sous-unités d’acide muramique. Lorsque leur proximité le permet, les chaînes tétrapeptidiques qui dépassent du squelette du glycane peuvent être réticulées par une liaison interpeptidique entre un groupe amino libre sur le DAP et un groupe carboxy libre sur un D-ala voisin.L’assemblage du peptidoglycane sur l’extérieur de la membrane plasmique est médié par un groupe d’enzymes périplasmiques, qui sont des transglycosylases, des transpeptidases et des carboxypeptidases. Le mécanisme d’action de la pénicilline et des bêta-lactamines apparentées consiste à bloquer les enzymes transpeptidases et carboxypeptidases pendant l’assemblage de la paroi cellulaire de la murène. Par conséquent, on dit que les antibiotiques bêtalactames « bloquent la synthèse de la paroi cellulaire » chez les bactéries.
L’épine dorsale glycanique de la molécule de peptidoglycane peut être coupée par une enzyme appelée lysozyme qui est présente dans le sérum animal, les tissus et les sécrétions, et dans le lysosome phagocytaire. La fonction du lysozyme est de lyser les cellules bactériennes, ce qui constitue une défense constitutive contre les agents pathogènes bactériens. Certaines bactéries à Gram positif sont très sensibles au lysozyme et l’enzyme est très active à de faibles concentrations.Les sécrétions lacrymales (larmes) peuvent être diluées au 1:40 000 et conserver la capacité de lyser certaines cellules bactériennes. Les bactéries Gram-négatives sont moins vulnérables à l’attaque du lysozyme car leur peptidoglycane est protégé par la membrane externe. Le site exact du clivage lysozymal est la liaison bêta 1,4 entre l’acide N-acétylmuramique (M) et la N-acétylglucosamine (G), de sorte que la sous-unité d’acideuramique représentée sur la figure 16(a) est le résultat de l’action du lysozyme sur le peptidoglycane bactérien.
Dans les bactéries à Gram positif, il existe de nombreux arrangements peptidiques différents parmi les peptidoglycanes. La plus étudiée est celle de Staphylococcus aureus représentée sur la figure 17 ci-dessous. A la place de la DAP (chez E. coli) se trouve le diaminoacide, la L-lysine (L-lys), et à la place de la liaison interpeptidique (chez les Gram-négatifs) se trouve un pont interpeptidique d’acides aminés qui relie un groupe amino libre sur la lysine à un groupe carboxy libre sur la D-ala d’une chaîne latérale tétrapeptidique voisine. Cet arrangement permet apparemment des liaisons croisées plus fréquentes entre les chaînes latérales tétrapeptidiques voisines. Chez S. aureus, le pont interpeptidique est un peptide composé de 5 molécules de glycine (appelé pont pentaglycine). L’assemblage du pont interpeptidique dans la muréine Gram-positive est inhibé par les antibiotiques bêta-lactames de la même manière que la liaison interpeptidique dans la muréine Gram-négative. Les bactéries à Gram positif sont plus sensibles à la pénicilline que les bactéries à Gram négatif car le peptidoglycane n’est pas protégé par une membrane externe et c’est une molécule plus abondante. Chez les bactéries à Gram positif, les peptidoglycanes peuvent varier en fonction de l’acide aminé qui remplace le DAP ou le L-lys en position 3 du tétrapeptide, et de la composition exacte du pont interpeptidique. Au moins huit types différents de peptidoglycanes existent chez les bactéries Gram-positives.
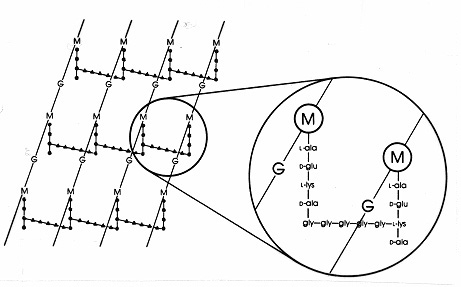
Figure 17. Schéma de la feuille de peptidoglycane de Staphylococcus aureus. G = N-acétyl-glucosamine ; M = acide N-acétyl-muramique ; L-ala = L-alanine ; D-ala = D-alanine ; D-glu = acide D-glutamique ; L-lys = L-lysine. Il s’agit d’un type de muréine que l’on trouve dans les bactéries Gram-positives. Par rapport au peptidoglycane d’E. coli (figure 7), la L-lys remplace le DAP (acide diaminopimélique) dans le tétrapeptide. Le groupe amino libre de la L-lys est remplacé par un glycinepentapeptide (gly-gly-gly-gly-gly-) qui devient alors un pont interpeptidique formant une liaison avec un groupe carboxy de la D-ala dans une chaîne latérale tétrapeptidique adjacente. Les peptidoglycanes à Gram positif diffèrent d’une espèce à l’autre, principalement en ce qui concerne les acides aminés en troisième position de la chaîne tétrapeptidique et la composition en acides aminés du pont interpeptidique.
Les bactéries à Gram négatif peuvent contenir une seule couche monomoléculaire de muréine dans leur paroi cellulaire, tandis que les bactéries à Gram positif sont censées avoir plusieurs couches ou « enveloppes » de peptidoglycane. Un groupe de molécules appelées acides teichoïques est étroitement associé aux couches de peptidoglycane des bactéries à Gram positif. Les acides teichoïques sont des polymères linéaires de polyglycérol ou de polyribitol substitués par des phosphates et quelques acides aminés et sucres. Les polymères d’acide teichoïque sont parfois ancrés à la membrane plasmique (appelés acide lipoteichoïque, LTA) et apparemment dirigés vers l’extérieur à angle droit par rapport aux couches de peptidoglycane. Les fonctions de l’acide téichoïque ne sont pas connues. Ils sont essentiels à la viabilité des bactéries Gram-positives dans la nature. Une idée est qu’ils fournissent un canal de charges négatives régulièrement orientées pour faire passer des substances chargées positivement à travers le réseau compliqué du peptidoglycane. Une autre théorie est que les acides teichoïques participent d’une manière ou d’une autre à la régulation et à l’assemblage des sous-unités d’acides muramiques à l’extérieur de la membrane plasmique. Il existe des cas,notamment chez les streptocoques, où les acides teichoïques ont été impliqués dans l’adhérence des bactéries aux surfaces des tissus.
chapitre suite
Page précédente
.
Laisser un commentaire