Semaine du requin : Que mangeait le mégalodon ?
On décembre 13, 2021 by adminAuteur invité : Jack Cooper, MSc
UoB Graduate / doctorant, Université de Swansea
Comme nous l’avons vu jusqu’à présent, il y a eu des changements et des mises à jour du consensus scientifique sur l’assignation taxonomique et la taille corporelle maximale de Megalodon. Cependant, sa collation préférée est quelque chose qui fait l’objet d’un consensus universel dans la communauté scientifique. Autre rareté, il s’agit d’une chose qui est généralement décrite avec une assez bonne précision dans la fiction. Permettez-moi de vous présenter un requin qui mangeait des baleines – de vraies baleines !
Plusieurs des grands requins macropédateurs actuels sont connus pour manger des mammifères marins, notamment notre vieil ami le grand requin blanc. Certaines des photographies les plus emblématiques de ce requin le montrent se jetant hors de l’eau avec sa dernière victime entre ses mâchoires (figure 1). Cependant, les jeunes requins blancs ne se nourrissent pas de phoques dès le début de leur vie. Ils commencent plutôt par manger des poissons et des requins plus petits. Si nous regardons à nouveau la formation de Gatun au Panama, nous trouvons beaucoup de requins fossiles, de raies et d’otolithes (structures de l’oreille interne des poissons), ainsi qu’une pléthore d’invertébrés comme des mollusques .
Ce que nous ne trouvons pas, cependant, ce sont des mammifères marins fossiles. Cela semble indiquer que les jeunes Megalodon mangeaient aussi des poissons et des requins plus petits plutôt que de s’attaquer immédiatement aux mammifères marins. C’est ce qu’on appelle un changement ontogénétique de régime alimentaire, c’est-à-dire que le requin change de source de nourriture préférée au cours de sa croissance. En mangeant des aliments plus gros, et probablement en grande quantité, lorsqu’il atteignait sa taille adulte, le Megalodon aurait été capable de maintenir les exigences métaboliques liées à sa taille. Il aurait pu le faire en mangeant non seulement des baleines, mais à peu près tout ce qu’il rencontrait.

Vu la quantité de nourriture que le Megalodon aurait dû manger pour justifier sa taille énorme, il était presque certainement un mangeur opportuniste. Le registre fossile est notre meilleur guide ici. Cela est dû à la présence de fossiles traces, qui sont des enregistrements géologiques de l’activité biologique. Il peut s’agir d’empreintes de dinosaures ou d’éraflures sur des os indiquant que quelque chose a mordu l’animal auquel l’os appartenait. Et le Megalodon a laissé quelques-unes de ces traces fossiles derrière lui, fournissant des preuves clés de ses meurtres.
Plusieurs fossiles ont été trouvés appartenant à une variété de cétacés avec des marques de morsure notables sur eux. Un exemple bien cité provient de la formation de Yorktown dans le Maryland, où plusieurs os de baleines présentaient des marques de morsure qui avaient clairement été faites par de grandes dents dentées . En fait, les dents de Megalodon sont parfois trouvées directement associées à ces os .
Les requins perdent et remplacent leurs dents des milliers de fois au cours de leur vie, et donc les requins perdent fréquemment des dents lors d’une mise à mort violente. C’est une raison essentielle pour laquelle les dents de requin sont des fossiles si communs. La présence de dents de requin à proximité d’un os de baleine rayé indique qu’une alimentation était en cours. Mais les paléontologues doivent toujours se poser la question suivante lorsque de tels fossiles sont découverts : s’agissait-il vraiment d’une prédation ? Ou était-ce peut-être du charognage ?
Le charognage est un comportement alimentaire courant chez les requins macropredateurs modernes tels que les grands blancs (Fig. 2) et les requins tigres . Lorsque ces requins rencontrent des baleines mortes flottant à la surface, c’est un buffet à volonté. Le charognage ne nécessite aucune dépense d’énergie, ce qui peut être gaspillé lors d’une tentative de prédation ratée. Étant donné que le Megalodon devait être capable de maintenir son énorme taille, il ne serait pas très logique de laisser passer un repas gratuit s’il rencontrait une baleine morte. Ainsi, par procuration de la façon dont les grands requins modernes se nourrissent, la plupart d’entre nous seraient prêts à parier que Megalodon a probablement fait du charognage lorsqu’il en avait l’occasion.

Cela dit, il existe un fossile particulièrement remarquable qui implique le Megalodon dans une tentative de prédation. Et, de manière cruciale, nous le savons principalement parce que sa tentative a échoué. Un article publié en 2010 décrit un fossile partiel de côte du Pliocène appartenant à un cétacé de la formation de Yorktown en Caroline du Nord (yep, deux formations de Yorktown). Ce fossile présentait également des dommages ressemblant à des marques de morsure, mais il semblait notamment qu’un os tissé recouvrait ces marques. Les auteurs ont interprété cela comme le signe d’une infection survenue pendant que l’animal se remettait du traumatisme osseux causé par ces marques de morsure. Ces marques correspondaient bien à une morsure dentelée, ce qui suggère que l’agresseur avait des dents dentelées – tout comme le Megalodon. L’étude a donc suggéré qu’un Megalodon, ou un autre grand requin, avait attaqué l’animal dont provenait le fossile, mais la victime n’avait survécu à la tentative que pour mourir d’une infection due à sa blessure six semaines plus tard .
Plus récemment encore, les toutes premières marques de morsure fossiles trouvées dans l’hémisphère sud attribuées à un Megalodon ont été découvertes dans la formation de Pisco au Pérou . Peut-être que cela devait être, car c’est le même site où Gordon Hubbell a trouvé son exceptionnel fossile de Carcharodon (discuté dans Q1). Le matériel mordu trouvé ici comprenait des restes de crâne et de côtes, suggérant que le requin avait mordu sa cible par le côté. Plus important encore, les restes ont été identifiés à une espèce individuelle pour la première fois – cette espèce étant Piscobalaena nana, une petite baleine à fanons. Comme pour les autres traces fossiles, les marques de morsure correspondent bien à de grandes dents dentelées – un bon argument pour accuser Megalodon de l’attaque (Fig. 3). Il est donc apparu que le Megalodon était également capable de cibler des baleines plus petites (qu’il s’agisse de prédation ou de charognage), ce qui est tout à fait conforme aux suggestions selon lesquelles il ciblait probablement aussi les dauphins, les tortues de mer et d’autres proies plus petites .

Si les stratégies de prédation sont difficiles à interpréter à partir de fossiles, les requins modernes peuvent nous donner une idée de la façon dont Megalodon chassait. La force de sa morsure aurait été exceptionnellement puissante pour endommager les os des baleines. Un modèle informatique du crâne d’un requin blanc a montré que sa force de morsure dépassait environ 1,8 tonne, ce qui en fait l’une des forces de morsure les plus puissantes de tous les animaux vivants. Par extension, cette étude suggère que la force de morsure de Megalodon était encore plus grande, estimée à un extraordinaire 108 514-182 201 N.
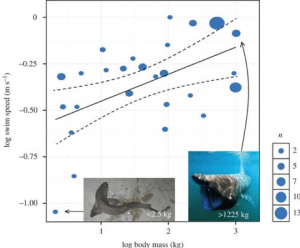
Les scientifiques ont également tenté de calculer la vitesse de nage de Megalodon, ce qui pourrait nous donner une indication de la façon dont il chassait. Les requins peuvent avoir une vitesse de rafale, c’est-à-dire un sursaut de nage rapide, généralement utilisé pour attraper une proie. Mais leur vitesse habituelle pour leurs nages occasionnelles est ce que nous appelons une vitesse de nage soutenue. Cela varie d’une espèce à l’autre, mais une étude dirigée par David Jacoby a combiné des données sur 26 espèces provenant de 64 études antérieures pour élaborer un modèle permettant de prédire une relation d’échelle entre la vitesse de nage soutenue et le métabolisme et la masse corporelle du requin. Leur modèle intégrant le métabolisme a révélé un exposant d’échelle de 0,173, résumé par l’équation suivante :
Vitesse de nage (ms ¯¹) ∝ Masse (kg)^0,173
Lorsqu’ils ont corrigé les données pour la phylogénie, cependant, ils ont constaté que les vitesses de nage minimales s’échelonnaient avec la masse corporelle à l’exposant légèrement inférieur de 0,15. L’équation ci-dessus suppose en outre une constante de 1 pour correspondre aux proportions, mais les données ont révélé que cette constante était en fait de 0,266. Ils ont donc construit l’équation de puissance :
Vitesse de nage (ms ¯¹) = 0,266 Masse (kg) ^0,15
À partir de là, ils sont capables de calculer la vitesse de nage soutenue du Megalodon en fonction de sa masse. Par exemple, le Megalodon de ~48 000 kg calculé comme la masse d’un requin de 15,9 m dans le travail de Gottfried révélerait un requin nageant continuellement à 1,34 m/s – environ 4,8 km/hr . L’application de ce modèle à un Megalodon de 52 000 kg révèle une vitesse de nage soutenue de 1,36 m/s, soit 4,9 km/h, tandis que si l’on allait vraiment à l’extrême et que l’on simulait un requin de 100 000 kg, ce modèle donnerait une vitesse de nage de 1,50 m/s, soit 5,4 km/h. Ce modèle s’est avéré très efficace non seulement pour calculer la vitesse de nage du Megalodon, mais aussi pour fournir un modèle pour de multiples espèces de requins (Fig. 4) .
Cependant, c’est la vitesse d’éclatement que les requins utilisent pour arracher rapidement leurs proies. Les requins blancs l’utilisent pour leur « comportement de brèche » dans lequel ils se propulsent hors de l’eau en se précipitant sur leur proie par le bas pour les prendre par surprise (Fig. 1) . Ils peuvent le faire car ils sont bien camouflés grâce à la contre-ombre. Donc, parce que le Megalodon était probablement contre-ombré aussi, et avait une écologie et une structure corporelle similaires à celles des requins blancs , cela pose la question : le Megalodon était-il aussi capable de faire du breaching ?
Peut-être. La vitesse d’éclatement du requin blanc est alimentée par une adaptation physiologique appelée mésothermie . C’est quelque chose que Megalodon a récemment été suggéré d’avoir également (à discuter en détail dans Q5). Les poissons mésothermes ont généralement des vitesses de nage soutenues et de rafale plus rapides que les poissons ectothermes , et cela a été utilisé pour suggérer que Megalodon pourrait avoir eu une vitesse de rafale allant jusqu’à 37,15 km/h .
Cette vitesse incroyablement rapide pour un si grand requin pourrait bien être suffisante pour être capable de faire une brèche. En outre, le requin pèlerin (Cetorhinus maximus), le deuxième plus grand requin actuel, a également été observé en train d’ouvrir une brèche . Peut-être le mégalodon géant était-il effectivement capable de bondir hors de l’eau pour attraper son dernier repas. Nous ne le savons pas encore, mais mon opinion personnelle et (quelque peu) professionnelle est que cela aurait bien pu être possible.
Avec un menu aussi large à choisir, le Megalodon était un prédateur apex, sans aucun prédateur connu en propre. Cela signifie deux choses importantes. Premièrement, il avait probablement une forte concurrence pour ses proies, ce qui a peut-être contribué à son extinction. Deuxièmement, en tant que prédateur apex, le Megalodon aurait été de la plus haute importance pour son écosystème, car il aurait gardé ses populations de proies sous contrôle et ainsi préservé un écosystème sain.
C’est pourquoi les requins d’aujourd’hui sont si importants pour les environnements marins. La perte d’un requin prédateur apex produit des effets en cascade irréversibles qui modifient la communauté qu’il laisse derrière lui . Ainsi, si nos écosystèmes marins sont ce qu’ils sont, c’est en partie parce que le Megalodon n’en fait plus partie. Et les énormes baleines des océans modernes peuvent dormir tranquilles en conséquence.
- Estrada JA, Rice AN, Natanson LJ & Skomal GB 2006. Utilisation de l’analyse isotopique des vertèbres pour reconstruire l’écologie alimentaire ontogénétique chez les requins blancs. Ecology 87, 829-834.
- Pimiento C, Ehret DJ, MacFadden BJ & Hubbell G 2010. Ancienne zone de nurserie pour le requin géant éteint Megalodon du Miocène du Panama. PLoS One 5, e10552.
- Pimiento C, González-Barba G, Ehret DJ, Hendy AJ, MacFadden BJ & Jaramillo C 2013. Requins et raies (Chondrichthyes, Elasmobranchii) de la formation Gatun du Miocène tardif du Panama. J. Paleontol. 87, 755-774.
- Purdy RW 1996. Paléoécologie des requins blancs fossiles. In : Grands requins blancs : la biologie de Carcharodon carcharias (eds. Klimley AP & Ainley DG), 67-78, Academic Press, San Diego.
- Aguilera OA, García L & Cozzuol MA 2008. Requins blancs à dents de géant et interaction trophique des cétacés de la Formation Paraguaná des Caraïbes du Pliocène. Paläontol. Z. 82, 204-208.
- Fallows C, Gallagher A. & Hammerschlag N 2013. Le charognard des requins blancs (Carcharodon carcharias) sur les baleines et son rôle potentiel pour façonner davantage l’écologie d’un prédateur apex. PLoS One, 8, e60797.
- Dudley SFJ, Anderson-Reade MD, Thompson GS & McMullen PB 2000. Récupération simultanée d’une carcasse de baleine par les grands requins blancs, Carcharodon carcharias, et les requins tigres, Galeocerdo cuvier. Fish. Bull. 98, 646-649.
- Kallal RJ, Godfrey SJ & Ortner DJ 2010. Les réactions osseuses sur une côte de cétacé du Pliocène indiquent la survie à court terme d’un événement de prédation. Int. J. Osteoarchaeol. 22, 253-260.
- Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M & Bianucci G 2017. Le requin géant éteint Carcharocles megalodon ciblait-il les petites proies ? Des marques de morsure sur des restes de mammifères marins de la fin du Miocène du Pérou. Palaeogeogr. Palaeoclimatol. Palaeoecol. 469, 84-91.
- Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, Ferrara TL, Cunningham E, Dean MN & Summers AP 2008. Analyse informatique tridimensionnelle de la mécanique de la mâchoire du requin blanc : à quel point un grand blanc peut-il mordre ? J. Zool. 276, 336-342.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2015. L’échelonnement de la vitesse de nage chez les requins est-il piloté par le métabolisme ? Biol. Lett. 11, 20150781.
- Gottfried MD, Compagno LJV & Bowman SC 1996. Taille et anatomie squelettique du requin géant « megatooth » Carcharodon megalodon. In : Grands requins blancs : la biologie de Carcharodon carcharias (eds. Klimley AP & Ainley DG), 55-66, Academic Press, San Diego.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2016. Correction à ‘Is the scaling of swim speed in sharks driven by metabolism?’. Biol. Lett. 12, 20160775.
- Watanabe YY, Payne NL, Semmens JM, Fox A & Huveneers C 2019. Stratégies de nage et énergétique des requins blancs endothermiques pendant la recherche de nourriture. J. Exp. Biol. 222, jeb185603.
- Cooper JA, Pimiento C, Ferrón HG & Benton MJ (sous presse). Dimensions du corps du requin géant éteint Otodus megalodon : Une reconstruction en 2D. Sci. Rep.
- Watanabe YY, Goldman KJ, Caselle JE, Chapman DD & Papastamatiou YP 2015. Les analyses comparatives des données de suivi des animaux révèlent l’importance écologique de l’endothermie chez les poissons. Proc. Nat. Acad. Sci. 112, 6104-6109.
- Ferrón HG 2017. L’endothermie régionale comme déclencheur du gigantisme chez certains requins macropredateurs éteints. PLoS One, 12, e0185185.
- Johnston EM, Halsey LG, Payne NL, Kock AA, Iosilevskii G, Whelan B &Houghton JD 2018. Puissance latente des requins pèlerins révélée par des événements exceptionnels de breaching. Biol. Lett. 14, 20180537.
- Myers RA, Baum JK, Shepherd TD, Powers SP & Peterson CH 2007. Effets en cascade de la perte de requins prédateurs apex d’un océan côtier. Science, 315, 1846-1850.
Éditée par Rhys Charles
.
Laisser un commentaire