Noyau olivaire supérieur
On novembre 12, 2021 by admin| Cerveau : Complexe olivaire supérieur | ||
|---|---|---|
 |
||
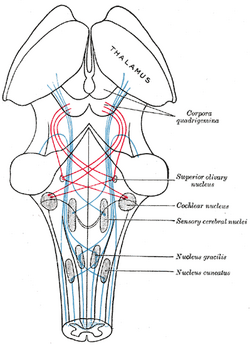
| Schéma montrant le parcours des fibres du lemniscus ; lemniscus médial en bleu, latéral en rouge. (Le noyau olivaire supérieur est étiqueté au centre droit.) | ||
| ] | ||
| Latin | nucléus olivaris supérieur | |
| Nucléus de Gray | sujet n°187 787 | |
| Partie de | ||
| Composantes | ||
| Artère | ||
| Vein | ||
| BrainInfo/UW | {{{BrainInfoType}}-.565 | |
| MeSH | ||
Le complexe olivaire supérieur (ou SOC ou olive supérieure) est une petite masse de substance grise située sur la face dorsale de la partie latérale du corps trapézoïdal. Petit chez l’homme, mais bien développé chez certains animaux, il présente une structure similaire à celle du noyau olivaire inférieur, et est situé immédiatement au-dessus de celui-ci. Certaines des fibres du corps trapézoïdal se terminent en s’arborant autour des cellules de ce noyau, tandis que d’autres naissent de ces cellules.
Physiologie
Le noyau olivaire supérieur joue un certain nombre de rôles dans l’audition. L’olive supérieure médiale (OSM) est un noyau spécialisé qui est censé mesurer la différence de temps d’arrivée des sons entre les oreilles (la différence de temps interaurale ou DTI). L’ITD est un indice majeur pour déterminer l’azimut des sons de basse fréquence, c’est-à-dire les localiser sur le plan azimutal – leur degré vers la gauche ou la droite.
L’olive latérale supérieure (LSO) serait impliquée dans la mesure de la différence de niveau des sons entre les oreilles (la différence de niveau interaurale ou ILD). L’ILD est un deuxième indice majeur dans la détermination de l’azimut des sons de haute fréquence.
Relation avec le système auditif
Le complexe olivaire supérieur est généralement situé dans le pons, mais chez l’homme, il s’étend de la médulla rostrale au mid-pons et reçoit des projections principalement du noyau cochléaire antéroventral via la strie acoustique ventrale, bien que le noyau postéroventral s’y projette également via la strie acoustique intermédiaire. C’est à cet endroit que se produisent les premières grandes interactions binaurales.
Noyaux primaires
Le complexe olivaire supérieur est divisé en trois noyaux primaires, le MSO, le LSO et le noyau médian du corps trapézoïdal, et plusieurs noyaux périolivaires plus petits. Ces trois noyaux sont les plus étudiés, et donc les mieux compris. Typiquement, ils sont considérés comme formant la voie de localisation azimutale ascendante.
Olivier supérieur médian (OSM)
On pense que l’olive supérieure médiane aide à localiser l’azimut d’un son, c’est-à-dire l’angle à gauche ou à droite où se trouve la source sonore. Le premier réflexe peut être de penser que ce noyau inclut des informations verticales, mais ce n’est pas le cas ; les informations traitées dans l’OSM sont limitées au plan horizontal. Les cellules fusiformes ne se projettent sur rien au niveau du pons, et n’interviennent qu’au niveau du colliculus inférieur. Seules des données horizontales sont présentes, mais elles proviennent de deux sources auditives différentes, ce qui facilite la localisation du son sur l’axe azimutal. Le colliculus supérieur y parvient en mesurant les différences de temps entre les signaux de deux oreilles enregistrant le même stimulus. Le déplacement autour de la tête prend environ 700 μs, et on suppose que l’olive supérieure médiane est capable de le détecter. En fait, on observe que les gens peuvent détecter des différences interaurales jusqu’à 10 microsecondes. Le noyau est organisé de manière tonotopique, mais la projection azimutale du champ réceptif est « très probablement une carte complexe et non linéaire. »
Les projections de l’olive supérieure médiane se terminent densément dans le noyau central ipsilatéral du colliculus inférieur. La majorité de ces axones sont considérés comme étant de « forme ronde » ou de type R. Ces axones R sont principalement glutamatergiques et contiennent des vésicules synaptiques rondes et forment des jonctions synaptiques asymétriques.
- C’est le plus grand des noyaux et chez l’homme, il contient environ 15 500 neurones .
- Chaque MSO reçoit des entrées bilatérales à basse fréquence des AVCN droit et gauche.
- La sortie se fait vers le lemniscus latéral ipsilatéral et finalement vers le colliculus inférieur.
- Le MSO répond mieux aux stimuli binauraux.
- Sa fonction principale implique la détection des différences de temps d’arrivée des sons aux deux oreilles, ce qui fait partie du processus de localisation (DTI).
L’olive latérale supérieure (LSO)
Cette olive a des fonctions similaires à l’olive médiale supérieure, mais emploie l’intensité pour localiser la source sonore. C’est la partie du tronc cérébral qui étiquette le son le plus fort provenant de l’oreille gauche comme étant du côté gauche. L’olive latérale reçoit des informations des deux noyaux cochléaires, des cellules sphériques buissonnantes de l’AVCN ipsilatéral et des cellules globulaires buissonnantes de l’AVCN controlatéral via le MNTB ipsilatéral. Les cellules principales du MNTB sont glycinergiques, fournissant une source d’inhibition à la LSO, et constituant la base de la sensibilité à l’ILD. Les projections des deux noyaux cochléaires sont principalement de haute fréquence, et ces fréquences sont ensuite représentées par la majorité des neurones du LSO (>2/3 sur 2-3kHz chez le chat). Des entrées supplémentaires proviennent du LNTB ipsilatéral (glycinergiques, voir ci-dessous), qui fournissent des informations inhibitrices du noyau cochléaire ipsilatéral. Une autre entrée possiblement inhibitrice provient des cellules non sphériques de l’AVCN ipsilatéral. Ces cellules sont soit globulaires touffues, soit multipolaires (stellaires). L’une ou l’autre de ces deux entrées pourrait fournir la base de l’inhibition ipsilatérale observée dans les cartes de réponse flanquant l’excitation primaire, affinant l’accord en fréquence de l’unité.
Le LSO se projette bilatéralement vers le noyau central du colliculus inférieur (ICC). Les projections ipsilatérales sont principalement inhibitrices (glycinergiques), et contralatérales excitatrices. Les cibles de projection supplémentaires comprennent les noyaux dorsal et ventral du lemniscus latéral (DNLL & VNLL). Les projections GABAergiques du DNLL constituent la principale source de GABA dans le tronc cérébral auditif, et se projettent bilatéralement vers l’ICC et le DNLL controlatéral. Ces connexions excitatrices et inhibitrices convergentes peuvent agir pour diminuer la dépendance au niveau de la sensibilité à la PEI dans la CCI par rapport à la DNL.
Des projections supplémentaires forment le faisceau olivocochléaire latéral (LOC), qui innerve les cellules ciliées internes cochléaires. On pense que ces projections ont une longue constante de temps, et agissent pour normaliser le niveau sonore détecté par chaque oreille afin d’aider à la localisation du son. Il existe des différences considérables entre les espèces : Les neurones de projection du LOC sont distribués à l’intérieur du LSO chez les rongeurs, et entourent le LSO chez les prédateurs (c’est-à-dire le chat).
Noyau médian du corps trapézoïdal (MNTB)
- Le MNTB est la plus petite partie du SOC ; la présence du MNTB dans le tronc cérébral humain est mise en doute.
- Chaque neurone ne reçoit qu’une entrée haute fréquence via un Calice de tenue de l’AVCN controlatéral, et sa sortie se projette vers le LSO ipsilatéral.
- On trouve deux types de réponse : un « type hacheur » semblable aux cellules fusiformes de l’AVCN et un type primaire qui ressemble à ceux des cellules buissonnières de l’AVCN.
Noyaux périolivaires
Le SOC est composé de six à neuf noyaux périolivaires, selon le chercheur cité, typiquement nommés en fonction de leur emplacement par rapport aux noyaux primaires. Ces noyaux entourent chacun des noyaux primaires et contribuent aux systèmes auditifs ascendants et descendants. Ces noyaux forment également la source du faisceau olivocochléaire, qui innerve la cochlée. Chez le cobaye, les projections ascendantes vers les colliculi inférieurs sont principalement ipsilatérales (>80%), la source unique la plus importante provenant du SPON. De même, les noyaux ventraux (RPO, VMPO, AVPO, & VNTB) sont presque entièrement ipsilatéraux, tandis que les autres noyaux se projettent bilatéralement.
| Nom | Chat | Cigarette | Rat | Souris | |
|---|---|---|---|---|---|
| LSO | X | X | X | X | |
| MSO | X | X | X | ||
| MNTB | X | X | X | X | X |
| LNTB | X | X | « LVPO » | X | |
| ALPO | X | X | |||
| PVPO | X | X | |||
| PPO | X | X | « CPO » | ||
| VLPO | X | ||||
| DPO | X | X | X | ||
| DLPO | X | X | |||
| VTB | X | X | « MVPO » | X | |
| AVPO | X | ||||
| VMPO | X | X | |||
| RPO | X | X | |||
| SPN | « DMPO » | X | X | X |
Nucléus ventral du corps trapézoïdal (VNTB)
- Le VNTB est un petit noyau situé latéralement au MNTB, et ventral au MSO.
- Composé d’une population hétérogène de cellules, ce noyau projette vers de nombreux noyaux auditifs, et forme le faisceau olivocochléaire médian (MOC) qui innerve les cellules ciliées externes cochléaires . Ces cellules contiennent des fibres électromotrices, et agissent comme des amplificateurs/atténuateurs mécaniques au sein de la cochlée.
- Le noyau se projette vers les deux CI, aucune cellule ne se projetant de manière bilatérale.
Noyau latéral du corps trapézoïdal (LNTB)
- Localisé ventralement au LSO
- Les cellules buissonnantes sphériques de l’AVCN projettent des collatérales bilatéralement, et les cellules buissonnantes globulaires projettent des collatérales ipsilatéralement aux neurones du LNTB.
- Les cellules sont immunoréactives pour la glycine, et sont marquées de façon rétrograde après injection de glycine tritiée dans le LSO
- Le noyau projette vers les deux IC, avec quelques cellules projetant de façon bilatérale, ainsi que vers le LSO ipsilatéral.
- De grandes cellules multipolaires projettent vers le noyau cochléaire, mais pas vers l’IC, chez le chat et le cobaye.
- Les entrées se font souvent par l’intermédiaire de bulbes terminaux tenus, produisant une transduction du signal très rapide.
Nucléus périolivaire supérieur (SPON) (Nucléus périolivaire dorsomédial (DMPO)).
- Situé directement en position dorsale par rapport au MNTB
- Chez le rat, le SPON est un noyau GABAergique homogène. Ces neurones organisés de façon tonotopique reçoivent des entrées excitatrices de la pieuvre et des cellules multipolaires du noyau cochléaire ventral controlatéral, une entrée glycinergique (inhibitrice) du MNTB ipsilatéral, une entrée GABAergique (inhibitrice) inconnue, et se projettent vers la CCI ipsilatérale. La plupart des neurones ne répondent qu’au décalage d’un stimulus, peuvent se verrouiller en phase sur des stimuli AM jusqu’à 200 Hz et peuvent constituer la base de la sélectivité de durée de la CCI. Notamment, les neurones SPON ne reçoivent pas d’entrées descendantes de la CCI, et il ne projette pas vers la cochlée ou le noyau cochléaire comme le font de nombreux noyaux périolivaires.
- Chez le cobaye, des cellules multipolaires rondes à ovales se projettent vers les deux CI, de nombreuses cellules se projetant de manière bilatérale. Les cellules plus allongées qui se projettent vers le noyau cochléaire ne se projettent pas vers le CCI. Il semble y avoir deux populations de cellules, l’une qui projette de manière ipsilatérale, et l’autre qui projette de manière bilatérale.
- La majorité des informations provenaient du SPON des rongeurs, en raison de la taille proéminente du noyau chez ces espèces, avec très peu d’études ont été faites sur le DMPO du chat, dont aucune n’était étendue.
Noyau périolivaire dorsal (DPO)
- Situé en position dorsale et médiale par rapport au LSO
- Contenant à la fois des unités EE (excitées par les deux oreilles) et E0 (excitées par l’oreille controlatérale uniquement).
- Les neurones sont organisés de façon tonotopique, et à haute fréquence.
- Peut appartenir à un seul noyau avec le DLPO
- Le noyau projette vers les deux CI, sans cellules se projetant bilatéralement.
Noyau périolivaire dorsolatéral (DLPO)
- Situé dorsalement et latéralement par rapport au LSO
- Contenant à la fois des unités EE (excitées par les deux oreilles) et E0 (excitées par l’oreille controlatérale seulement).
- Les neurones sont organisés de façon tonotopique, et de basse fréquence.
- Peut appartenir à un seul noyau avec le DPO
- Le noyau se projette vers les deux CI, avec peu de cellules se projetant bilatéralement.
Noyau périolivaire ventrolatéral (OPVL)
- Situé ventralement à et dans le hillus ventral du LSO
- Comprend à la fois des unités EI (excitées par l’oreille controlatérale et inhibées par l’oreille ipsilatérale) et E0 (excitées uniquement par l’oreille controlatérale).
- Les neurones sont organisés de façon tonotopique, et de haute fréquence.
Nucléus périolivaire antérolatéral (ALPO)
- Le noyau se projette vers les deux CI, sans cellules se projetant bilatéralement.
- De grandes cellules multipolaires se projettent sur le noyau cochléaire, mais pas sur le CI, chez le chat et le cobaye.
Noyau périolivaire ventromédian (VMPO)
- Situé entre le MSO et le MNTB.
- Envoie des projections vers l’ICC de manière bilatérale.
- Le noyau projette vers les deux CCI, aucune cellule ne se projetant de manière bilatérale.
Nucléus périolivaire rostral (NPR) (Nucléus périolivaire antérieur (NPA)).
- Situé entre le pôle rostral du MSO et le VNLL
Nucléus périolivaire caudal (CPO) (Nucléus périolivaire postérieur (PPO)).
- Situé entre le pôle caudal du MSO et le noyau facial (7N)
Noyau périolivaire postéroventral (PVPO)
- Le noyau se projette vers les deux CI, sans qu’aucune cellule ne se projette bilatéralement.
Physiopathologie
L’autopsie d’une femme de 21 ans atteinte d’autisme, d’épilepsie et de retard mental a révélé une absence quasi-complète de l’olive supérieure.
Voir aussi
- Corps olivaire
- Kulesza RJ, Cytoarchitecture du complexe olivaire supérieur humain : olive supérieure médiale et latérale. Hearing Research 225(2007) 80-90
- 2,0 2,1 Oliver DL, et al, Axonal projections from the lateral and medial superior olive to the inferior colliculus of the cat : a study using electron microscopic autoradiography. J Comp Neurol. 1995 Sep 11;360(1):17-32 Erreur de citation : Balise
<ref>non valide ; nom « R4 » défini plusieurs fois avec un contenu différent - Cajal, S. R. Y. et L. Azoulay (1909). Histologie du système nerveux de l’homme et des vertébrés. Paris, Maloine.
- 4.0 4.1 Kandel, et al Principles of Neuroscience. Quatrième édition. pp 591-624. Copyright 2000, par McGraw-Hill Co. Erreur de citation : Balise
<ref>non valide ; nom « R2 » défini plusieurs fois avec un contenu différent - Kulesza, R. J., Jr. (2007). « Cytoarchitecture du complexe olivaire supérieur humain : olive supérieure médiale et latérale ». Hear Res 225(1-2) : 80-90.
- Tsuchitani, C. et J. C. Boudreau (1967). « Encodage de la fréquence et de l’intensité du stimulus par les cellules du segment S de l’olive supérieure du chat ». J Acoust Soc Am 42(4) : 794-805.
- 8.0 8.1 8.2 Erreur de citation : Balise
<ref>non valide ; nom « R10 » défini plusieurs fois avec un contenu différent - Wu, S. H. et J. B. Kelly (1994). « Preuve physiologique de l’inhibition ipsilatérale dans l’olive supérieure latérale : réponses synaptiques dans une tranche de cerveau de souris ». Hear Res 73(1) : 57-64.
- Brownell, W. E., P. B. Manis, et al. (1979). « Réponses inhibitrices ipsilatérales dans l’olive supérieure latérale du chat ». Brain Res 177(1) : 189-93.
- Darrow, K. N., S. F. Maison, et al. (2006). « La rétroaction efférente cochléaire équilibre la sensibilité interaurale ». Nat Neurosci 9(12) : 1474-6.
- Warr, W. B. et J. J. Guinan, Jr. (1979). « Innervation efférente de l’organe de corti : deux systèmes distincts ». Brain Res 173(1) : 152-5.
- 13.0 13.1 Schofield, B. R. et N. B. Cant (1991). » Organisation du complexe olivaire supérieur chez le cobaye. I. Cytoarchitecture, histochimie de la cytochrome oxydase, et morphologie dendritique ». J Comp Neurol 314(4) : 645-70. Erreur de citation : Invalid
<ref>tag ; name « R7 » defined multiple times with different content - 14.0 14.1 14.2 14.3 14.4 14.5 14.6 14.7 14.8 Illing, R. B., K. S. Kraus, et al. (2000). « Plasticité du complexe olivaire supérieur ». Microsc Res Tech 51(4) : 364-81. Erreur de citation : Invalid
<ref>tag ; name « R5 » defined multiple times with different content Cite error : Invalid<ref>tag ; name « R5 » defined multiple times with different content Cite error : Balise<ref>non valide ; nom « R5 » défini plusieurs fois avec un contenu différent Erreur de citation : Balise<ref>non valide ; nom « R5 » défini plusieurs fois avec un contenu différent Erreur de citation : Balise<ref>non valide ; nom « R5 » défini plusieurs fois avec un contenu différent Erreur de citation : Balise<ref>non valide ; nom « R5 » défini plusieurs fois avec un contenu différent Erreur de citation : Balise<ref>non valide ; nom « R5 » défini plusieurs fois avec un contenu différent Erreur de citation : Invalid<ref>tag ; name « R5 » defined multiple times with different content - Warr, W. B. and J. E. Beck (1996). « Projections multiples du noyau ventral du corps trapézoïdal chez le rat ». Hear Res 93(1-2) : 83-101.
- 16.00 16.01 16.02 16.03 16.04 16.05 16.06 16.07 16.08 16.09 16.10 Schofield, B. R. et N. B. Cant (1992). » Organisation du complexe olivaire supérieur chez le cobaye : II. Patterns of projection from the periolivary nuclei to the inferior colliculus ». J Comp Neurol 317(4) : 438-55. Erreur de citation : Invalid
<ref>tag ; name « R6 » defined multiple times with different content Cite error : Invalid<ref>tag ; name « R6 » defined multiple times with different content Cite error : Balise<ref>non valide ; nom « R6 » défini plusieurs fois avec un contenu différent Cite error : Balise<ref>non valide ; nom « R6 » défini plusieurs fois avec un contenu différent Cite error : Balise<ref>non valide ; nom « R6 » défini plusieurs fois avec un contenu différent Cite error : Balise<ref>non valide ; nom « R6 » défini plusieurs fois avec un contenu différent Cite error : Balise<ref>non valide ; nom « R6 » défini plusieurs fois avec un contenu différent Cite error : Balise<ref>non valide ; nom « R6 » défini plusieurs fois avec un contenu différent Cite error : Balise<ref>non valide ; nom « R6 » défini plusieurs fois avec un contenu différent Cite error : Invalid<ref>tag ; name « R6 » defined multiple times with different content - Smith, P. H., P. X. Joris, et al. (1993). « Projections d’axones de cellules buissonnantes sphériques physiologiquement caractérisées à partir du noyau cochléaire du chat : preuve de l’existence de lignes de retard vers l’olive supérieure médiane ». J Comp Neurol 331(2) : 245-60.
- Wenthold, R. J., D. Huie, et al. (1987). « Immunoréactivité de la glycine localisée dans le noyau cochléaire et le complexe olivaire supérieur ». Neuroscience 22(3) : 897-912.
- 19.0 19.1 Adams, J. C. (1983). « Cytologie des cellules périolivaires et organisation de leurs projections chez le chat ». J Comp Neurol 215(3) : 275-89. Erreur de citation : Balise
<ref>non valide ; nom « R8 » défini plusieurs fois avec un contenu différent - Friauf, E. et J. Ostwald (1988). « Projections divergentes des neurones du noyau cochléaire ventral de rat physiologiquement caractérisés, comme le montre l’injection intra-axonale de peroxydase de raifort. » Exp Brain Res 73(2) : 263-84.
- Kulesza, R. J., Jr. et A. S. Berrebi (2000). « Le noyau paraolivaire supérieur du rat est un noyau GABAergique ». J Assoc Res Otolaryngol 1(4) : 255-69.
- Kulesza, R. J., Jr., G. A. Spirou, et al. (2003). « Propriétés de réponse physiologique des neurones du noyau paraolivaire supérieur du rat ». J Neurophysiol 89(4) : 2299-312.
- BLANC JS, WARR WB. La double origine du faisceau olivocochléaire chez le rat albinos. J. Comp. Neurol. 219:203-214, 1983.
- Guinan, J. J., Jr., S. S. Guinan, et al. (1972). « Unités auditives uniques dans le complexe olivaire supérieur. I. Réponses aux sons et classifications basées sur les propriétés physiologiques ». Int J Neurosci 4 : 101-20.
- Tsuchitani, C. (1977). « Organisation fonctionnelle des groupes de cellules latérales du complexe olivaire supérieur du chat ». J Neurophysiol 40(2) : 296-318.
- Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996). Origine embryologique de l’autisme : anomalies de développement des noyaux moteurs des nerfs crâniens. J Comp Neurol 370 (2) : 247-61.
- MedEd at Loyola Neuro/frames/nlBSs/nl27fr.htm
- Image at med.utah.edu (pick 9b. Pons)
Cet article était initialement basé sur une entrée d’une édition du domaine public de Gray’s Anatomy. En tant que tel, certaines des informations qu’il contient peuvent être dépassées. Veuillez éditer l’article si c’est le cas, et n’hésitez pas à supprimer cet avis lorsqu’il n’est plus pertinent.
|
v-d-e
Cerveau : rhombencéphale (cerveau postérieur) |
||||||
|---|---|---|---|---|---|---|
| Métencéphale/ pons |
|
Dorsal/
(tegmentum) |
Ventral/
(base) |
Raphe/reticulaire
|
||
| Myélencéphale/ médulla |
|
Surface dorsale
|
Ventral
|
Raphe/réticulaire
|
||
|
v-d-e
Système sensoriel : Systèmes auditif et vestibulaire (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oreille externe |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Earlobe) – Canal auditif – Muscles auriculaires Eardrum (Umbo, Pars flaccida) |
|||||||||||
| Moyenne oreille |
|
|||||||||||
| Oreille interne/ (labyrinthe membraneux, labyrinthe osseux) |
|
|||||||||||
| {| class= »navbox collapsible nowraplinks » style= »margin :auto ; » | ||||||||||||
|
—
|
||||||||||||
|
||||||||||||
|}
Cette page utilise un contenu sous licence Creative Commons de Wikipedia (voir les auteurs).
Laisser un commentaire