Frontières en Psychologie
On septembre 28, 2021 by adminLe corps calleux (CC) est la principale commissure reliant les deux hémisphères cérébraux du cerveau. Par conséquent, bien que le terme commissurotomie ne soit pas incorrect pour décrire le soi-disant « cerveau divisé », et qu’il soit strictement correct pour les opérations chirurgicales humaines originales dans lesquelles toutes les commissures ont été sectionnées (Gazzaniga et al., 1967), les chirurgies ultérieures n’ont sectionné que la CC, faisant ainsi entrer en usage le terme « callosotomie » qui reste comme le terme le plus approprié dans ces cas (par exemple, Franz et al., 1996, 2000).
D’un point de vue évolutif, la CC est un développement récent observé uniquement chez les mammifères placentaires (Katz et al., 1983 ; Mihrshahi, 2006). On pense que la fonctionnalité fournie par la CC a été précédée, au moins en partie, par la commissure antérieure (CA), comme on l’a observé chez les mammifères non placentaires tels que les marsupiaux (Heath et Jones, 1971 ; Ashwell et al., 1996). Bien que certaines voies résiduelles de la CA soient encore présentes dans le cerveau humain, sa connectivité et sa structure restent peu étudiées dans la littérature actuelle et les rares résultats sont plutôt incohérents (Lasco et al., 2002 ; Patel et al., 2010 ; Choi et al., 2011). Par exemple, les résultats basés sur l’imagerie par résonance magnétique (IRM) sont mitigés quant à savoir si le CA est de taille similaire chez les femmes et les hommes (Lasco et al., 2002 ; Patel et al., 2010). Dans une étude relativement récente utilisant la technique in-vivo d’imagerie du tenseur de diffusion (DTI), qui s’appuie sur la diffusion des molécules d’eau dans les axones, Patel et al. (2010) ont étudié huit sujets et ont réussi à isoler des fibres du CA chez cinq d’entre eux. Parmi les résultats, chez un sujet, une grande proportion de fibres de la branche postérieure du CA se déplaçait bilatéralement dans les régions pariétales où la plupart d’entre elles se terminaient, avec un plus petit faisceau se terminant dans le gyrus précentral, et encore un autre faisceau entrant dans le lobe temporal. De telles différences individuelles ont été constatées chez tous les participants, mais la majorité des sujets ont montré des projections bilatérales vers les cortex orbitofrontal, pariétal et temporal, vers lesquels le CC ne se projette pas clairement. Ensemble, les résultats impliquent que l’AC a des projections à la fois primaires et vestigiales vers les zones néocorticales où le CC ne se projette pas, comme d’autres l’ont également suggéré (Di Virgilio et al., 1999 ; Patel et al., 2010).
Selon nous, l’AAA s’appuie fortement sur les réseaux néocorticaux et la connectivité avec les structures sous-corticales telles que les ganglions de la base et le thalamus (parmi d’autres non discutées spécifiquement dans notre proposition) (Franz, 2012). Ainsi, tout développement évolutif impliquant le CA pourrait également avoir un certain rôle dans le modèle AAA. Cependant, nous avons trouvé peu de littérature traitant spécifiquement de ce lien possible.
Le rôle de l’AC a été étudié dans des tâches qui sont susceptibles de nécessiter des formes d’attention, comme celles impliquant l’attention visuelle. Par exemple, il est bien connu que les populations acallosales ont été étudiées dans le contexte de tâches impliquant l’intégration visuelle (Gazzaniga et al., 1962, 1965 ; Sperry, 1968 ; Levy et al., 1972). Plus récemment, Corballis (1995) a cherché à aborder l’intégration visuelle et l’attention dans le but d’expliquer les divergences antérieures dans la recherche sur l’acallosal. Corballis a proposé un système d’attention visuelle dichotomique ; une partie (impliquant la forme) est automatique et localisée dans chaque hémisphère, et l’autre (impliquant le mouvement et la localisation) est volontaire et sous-corticale. Le modèle AAA peut s’appuyer sur cette observation et répondre aux divergences observées dans la littérature. D’après le modèle que nous proposons, à travers les multiples niveaux d’attention, il existe des voies autres que le CC qui pourraient servir de médiateur au transfert d’informations entre les hémisphères. Une possibilité implique le circuit des ganglions de la base qui, selon nous, est potentiellement critique dans la médiation du partage de l’attention entre les hémisphères (Franz, 2012). En transectant le CC, le transfert direct de l’attention par les voies corticales est largement aboli (à moins, bien sûr, que le CA ne joue un rôle de secours). Pourtant, l’intégration de l’information visuelle basée sur l’attention pourrait toujours être médiée par des voies sous-corticales et/ou le CA. De cette manière, le système d’attention en deux parties observé par Corballis (1995) pourrait être présent en raison de la médiation basée sur l’attention de l’intégration visuelle via l’AC (et/ou les ganglions de la base et les circuits impliqués : Franz, 2012).
Les études impliquant la vision et l’attention ne doivent pas être considérées entièrement séparément du système régulant l’action motrice. Sur la base de nos propres résultats (Franz, 2004 ; Franz et Packman, 2004), diverses formes d’attention (visuelle et interne-non visuelle) sont étroitement liées à l’action motrice, et il va de soi qu’une clé possible pour comprendre un rôle potentiel d’attention de l’AC réside dans le lien entre l’action, la vision et l’attention. Dans une étude antérieure, Franz (2004) a demandé à des participants normaux de dessiner des cercles de façon bimanuelle sous différentes manipulations du feedback visuel ou de l’attention interne à chaque main. La taille des cercles dessinés par une main recevant une forme d’attention interne est devenue plus grande d’une manière similaire aux effets qui se sont produits avec la présence d’un retour visuel de cette main (par rapport à l’absence d’attention/retour visuel). Notre modèle AAA implique que l’attention est un élément clé dans la régulation de la sortie motrice (ainsi que dans la sélection de l’action motrice planifiée), et dans la mesure des preuves actuelles, nous ne pouvons pas exclure que l’AC joue un rôle dans cette fonction. Cependant, le test direct du rôle du CA dans toute tâche cognitive n’est pas trivial.
Tester si le CA prend part à la fonction cognitive est presque impossible dans la recherche neuropsychologique étant donné que les participants avec un CA localisé ne sont pas faciles à trouver (et nous n’en connaissons aucun). Il pourrait donc être beaucoup plus intuitif d’évaluer les différences neurologiques dans le CA normal et de corréler les données neurologiques avec les données comportementales. La méthode de tractographie présentée (élucidation du CA) peut être appliquée relativement facilement à de larges échantillons de la population normale et, idéalement, peut être combinée avec les performances (basées sur des tâches effectuées au cours de sessions de test séparées de la collection DTI) de ces mêmes individus en utilisant des tâches cognitives conçues à cet effet. Nous démontrons ici que l’ITD peut être réalisée et donner des résultats utilisables sur un petit échantillon de personnes issues de la population neurologique normale. La proposition générale serait de mener des méthodes similaires sur un échantillon de plus grande taille, et (comme suggéré ci-dessus), de corréler les résultats (d’une mesure proxy de la densité de l’AC, par exemple) avec les variables d’une tâche cognitive.
Dans la figure 1, nous montrons les résultats du suivi de l’ITD sur les données obtenues auprès de 10 participantes provenant d’un ensemble de données publiques d’ITD à l’Université Johns Hopkins (voir Remerciements pour la source complète). L’âge moyen était de 22 ans (écart-type = 2,5). Les images ont été traitées à l’aide de la FSL Diffusion Toolbox (Smith et al., 2004 ; Woolrich et al., 2009 ; Jenkinson et al., 2012) et les procédures employées sont celles utilisées dans les études précédentes sur les autres trajets de fibres (FDT ; Behrens et al., 2003a,b, 2007 ; Johansen-Berg et al., 2004). En bref, les images ont d’abord été concaténées en volumes 4d pour chaque participant et les crânes ont été dépouillés à l’aide du Brain Extraction Tool (BET ; Smith, 2002 ; Jenkinson et al., 2005) de FSL. Les images extraites ont été corrigées pour tenir compte des mouvements mineurs de la tête et des courants de Foucault générés par les changements de direction du champ. Les tenseurs de diffusion ont été ajustés pour générer des images qui affichent les directions de diffusion primaires et les niveaux de FA à chaque voxel, à utiliser pour les cartes de couleur pendant le masquage. Une estimation bayésienne des paramètres de diffusion obtenus à l’aide de techniques d’échantillonnage avec des fibres croisées (BEDPOSTX ; « X » impliquant des fibres croisées) a été réalisée pour générer une distribution de la taille et de la direction de la diffusion tenant compte de deux fibres croisées afin de réaliser ultérieurement une tractographie à travers les voxels de diffusion.
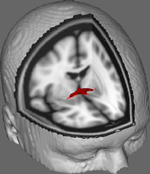
Figure 1. Résultats de suivi moyennés chez les 10 participants témoins superposés sur une image IRM standard pondérée en T1 avec des voxels isotropes de 2 mm pour la référence anatomique.
Les résultats de suivi moyennés chez les 10 participants témoins ont ensuite été superposés sur une image IRM standard pondérée en T1 avec des voxels isotropes de 2 mm pour la référence anatomique. Le rapport FA moyen du groupe était de 0,3146 (SD = 0,0472). Ces résultats de tractographie révèlent, de manière importante, que la structure de l’AC peut être élucidée, et a donc un potentiel pour d’autres études corrélant les scores de performance sur les tâches comportementales avec les valeurs FA en utilisant une plus grande taille d’échantillon.
En résumé, à l’heure actuelle, une littérature clairsemée existe, et parmi la littérature disponible, aucune ne relie spécifiquement l’AC aux processus liés au modèle AAA qui est le point central de notre article cible précédent (Franz, 2012). En utilisant une procédure similaire à celle proposée ici, il est possible de tester des échantillons plus importants de la population normale, et de corréler les valeurs FA des résultats DTI (par exemple) avec une variable dépendante spécifique obtenue à partir d’une tâche d’attention à l’action développée dans un but précis (dont les procédures sont un développement en cours dans notre laboratoire).
Déclaration de conflit d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de toute relation commerciale ou financière qui pourrait être interprétée comme un conflit d’intérêts potentiel.
Remerciements
Nous remercions sincèrement le Human Brain Project et le National Research Resource Center (USA) et l’Université John Hopkins pour avoir autorisé l’utilisation publique de leurs données DTI. Nous sommes également reconnaissants pour le généreux soutien financier du Marsden Fund de la Royal Society of New Zealand (à Elizabeth A. Franz).
Corballis, M. (1995). L’intégration visuelle dans le cerveau divisé. Neuropsychologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). L’allocation de l’attention à l’apprentissage d’actions dirigées vers un but : un cadre de neurosciences cognitives axé sur les ganglions de la base. Front. Psychol. 3:535. doi : 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., et Jones, E. (1971). Voies interhémisphériques en l’absence d’un corps calleux. Une étude expérimentale des connexions commissurales chez le phalanger marsupial. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
.
Laisser un commentaire