Superior olivary nucleus
On 12 marraskuun, 2021 by admin| Aivot: Superior olivary complex | ||
|---|---|---|
 |
||
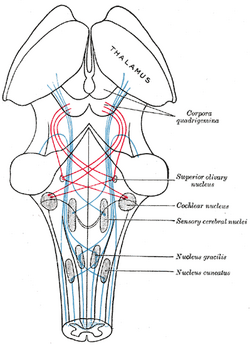
| Kaavio, jossa näkyy lemniscuksen kuitujen kulku; mediaalinen lemniscus sinisellä, lateraalinen punaisella. (Superior olivary nucleus on merkitty keskellä oikealla.) | ||
| ] | ||
| Latina | nucleus olivaris superior | |
| Grayn | subjektin #187 787 | |
| Taivutin | ||
| Komponentit | ||
| Arteria | ||
| Vein | ||
| Aivotieto/UW | {{{Aivotietotyyppi}}-565 | |
| MeSH | ||
Ylempi oliivikompleksi (superior olivary complex, SOC tai superior olive) on pieni harmaan aineen massa, joka sijaitsee trapetsirungon lateraalisen osan dorsaalipinnalla. Se on pieni ihmisillä, mutta hyvin kehittynyt tietyillä eläimillä, ja se on rakenteeltaan samanlainen kuin alempi olivariydin, ja se sijaitsee välittömästi sen yläpuolella. Osa trapetsirungon kuiduista päättyy arborisoitumalla tämän ytimen solujen ympärille, kun taas toiset lähtevät näistä soluista.
Fysiologia
Ylempänä olevalla olivariytimellä on useita tehtäviä kuulon kannalta. Mediaalinen ylempi oliivin ydin (MSO) on erikoistunut ydin, jonka uskotaan mittaavan äänien saapumisen aikaeroa korvien välillä (interauraalinen aikaero eli ITD). ITD on tärkeä vihje matalataajuisten äänien atsimuutin määrittämisessä eli niiden paikantamisessa atsimutaalitasolle – niiden astetta vasemmalle tai oikealle.
Lateraalisen ylemmän oliivin (lateral superior olive, LSO) uskotaan osallistuvan äänien tasoeron mittaamiseen korvien välillä (interauraalinen tasoero eli ILD). ILD on toinen tärkeä vihje korkeataajuisten äänien atsimuutin määrittämisessä.
Suhde kuulojärjestelmään
Ylempi oliivikompleksi sijaitsee yleensä ponsissa, mutta ihmisellä se ulottuu rostraalisesta medullasta keskiponssiin ja saa projisointeja pääasiassa anteroventraalisesta sisäkorvaperäisestä ytimestä ventraalisen akustisen säikeen välityksellä, joskin sinne projisoituu myös posteroventraalinen ydin väliakustisen säikeen välityksellä. Tässä kohdassa tapahtuvat ensimmäiset merkittävät binauraaliset vuorovaikutukset.
Primaariset ytimet
Ylempi olivariuskompleksi jakautuu kolmeen primaariseen ytimeen, MSO:een, LSO:een ja trapetsirungon mediaaliseen ytimeen, sekä useisiin pienempiin periolivariusytimiin. Nämä kolme ydintä ovat eniten tutkittuja ja siksi parhaiten ymmärrettyjä. Tyypillisesti niiden katsotaan muodostavan nousevan atsimutaalisen paikannusreitin.
Mediaalinen ylempi oliivi (MSO)
Mediaalisen ylemmän oliivin ajatellaan auttavan paikantamaan äänen atsimuutin eli sen kulman vasemmalle tai oikealle, jossa äänilähde sijaitsee. Ensivaisto saattaa ajatella, että tämä ydin sisältää vertikaalista tietoa, mutta näin ei ole; MSO:ssa käsiteltävä tieto rajoittuu horisontaaliseen tasoon. Fusiformiset solut eivät projisoi mihinkään ponsin tasolle, vaan ne tulevat mukaan toimintaan vasta colliculus inferiorissa. Ainoastaan horisontaalinen tieto on läsnä, mutta se tulee kahdesta eri korvalähteestä, mikä auttaa äänen paikallistamisessa atsimuuttiakselilla. Ylempi oliivi tekee tämän mittaamalla samaa ärsykettä tallentavien kahden korvasignaalin välisiä aikaeroja. Pään kiertäminen kestää noin 700 μs, ja oletetaan, että mediaalinen superior olive pystyy havaitsemaan tämän. Itse asiassa on havaittu, että ihmiset pystyvät havaitsemaan jopa 10 mikrosekunnin eroja. Ydin on tonotopisesti järjestäytynyt, mutta atsimutaalinen reseptiivisen kentän projektio on ”todennäköisesti monimutkainen, epälineaarinen kartta”.
Mediaalisen ylemmän oliivin projektiot päättyvät tiheästi colliculus inferiorin ipsilateraaliseen keskusytimeen. Suurinta osaa näistä aksoneista pidetään ”pyöreän muotoisina” eli tyypin R aksoneina. Nämä R-aksonit ovat enimmäkseen glutamatergisia ja sisältävät pyöreitä synaptisia vesikkeleitä ja muodostavat epäsymmetrisiä synaptisia liitoksia.
- Tämä on ytimistä suurin, ja ihmisellä se sisältää noin 15 500 neuronia.
- Kumpikin MSO saa matalataajuisia kahdenvälisiä syötteitä oikeasta ja vasemmasta AVCN:stä.
- Lähdöt suuntautuvat ipsilateraaliseen lateraaliseen lemniscukseen ja lopulta colliculus inferioriin.
- MSO vastaa paremmin binauraalisiin ärsykkeisiin.
- Sen päätehtävänä on havaita erot äänien saapumisajoissa molempiin korviin, mikä on osa lokalisointiprosessia (ITD).
Lateraalinen ylempi oliivi (LSO)
Tällä oliivilla on samanlaiset toiminnot kuin mediaalisella ylempänä olevalla oliivilla, mutta se käyttää intensiteettiä äänilähteen lokalisointiin. Tämä on aivorungon osa, joka leimaa vasemmasta korvasta tulevan kovemman äänen olevan vasemmalla puolella. Lateraalinen oliivi saa syötteitä molemmista sisäkorvan ytimistä, ipsilateraalisista AVCN:n pallomaisista puskasoluista ja kontralateraalisista AVCN:n pallomaisista puskasoluista ipsilateraalisen MNTB:n kautta. MNTB:n pääsolut ovat glyserergisiä, ne tarjoavat LSO:lle estolähteen ja muodostavat perustan ILD-herkkyydelle. Molemmista sisäkorvan ytimistä tulevat projektiot ovat pääasiassa korkeataajuisia, ja näitä taajuuksia edustaa myöhemmin suurin osa LSO:n neuroneista (>2/3 yli 2-3 kHz:n taajuudella kissalla). Lisäsyötteet tulevat ipsilateraalisesta LNTB:stä (glyserergiset, ks. jäljempänä), jotka antavat inhiboivaa tietoa ipsilateraalisesta sisäkorvan ytimestä. Toinen mahdollisesti inhiboiva tulo tulee ipsilateraalisista AVCN:n pallomaisista soluista. Nämä solut ovat joko pallomaisia puskasoluja tai monipolaarisia (tähtimäisiä). Kumpi tahansa näistä kahdesta syötteestä voisi muodostaa perustan ipsilateraaliselle inhibitiolle, joka näkyy primaarista herätettä reunustavissa vastekartoissa ja terävöittää yksikön taajuusviritystä.
LSO projisoituu bilateraalisesti colliculus inferiorin keskusytimeen (ICC). Ipsilateraaliset projektiot ovat ensisijaisesti inhiboivia (glyserergisiä) ja kontralateraaliset eksitatorisia. Muita projektiokohteita ovat lateraalisen lemniscuksen dorsaalinen ja ventraalinen ydin (DNLL & VNLL). DNLL:stä lähtevät GABAergiset projektiot muodostavat GABA:n ensisijaisen lähteen kuuloaivorungossa, ja ne projisoituvat bilateraalisesti ICC:hen ja kontralateraaliseen DNLL:ään. Nämä toisiaan lähentävät eksitatoriset ja inhibitoriset yhteydet saattavat vähentää ILD-herkkyyden tasoriippuvuutta ICC:ssä verrattuna LSO:han.
Lisäprojektiot muodostavat lateraalisen olivokokleaarisen kimpun (Lateral Olivocochlear Bundle, LOC), joka innervoi sisäkorvan sisempiä karvasoluja. Näillä projektioilla uskotaan olevan pitkä aikavakio, ja ne toimivat normalisoidakseen kummankin korvan havaitseman äänitason äänen paikallistamisen helpottamiseksi. Lajeissa on huomattavia eroja: Jyrsijöillä LOC-projektioneuronit ovat jakautuneet LSO:n sisälle ja ympäröivät LSO:ta petoeläimillä (esim. kissa).
Medial Nucleus of Trapezoid Body (MNTB)
- MNTB on SOC:n pienin osa; MNTB:n esiintyminen ihmisen aivorungossa on kyseenalaista.
- Jokainen neuroni saa vain korkeataajuista tuloa kontralateraaliselta AVCN:ltä pidetyn calyxin kautta, ja sen ulostulo projisoituu ipsilateraaliseen LSO:han.
- Havaitaan kaksi vastetyyppiä: AVCN:n karasolujen kaltainen ”chopper-tyyppi” ja AVCN:n puskasolujen kaltainen primäärityyppi.
Periolivary Nuclei
SOC koostuu kuudesta – yhdeksästä periolivaryntäisestä ytimestä riippuen siteeratusta tutkijasta, ja ne on tyypillisesti nimetty sijaintinsa perusteella primaarisiin ytimiin verrattuna. Nämä ytimet ympäröivät kutakin primaarista ydintä, ja ne vaikuttavat sekä nousevaan että laskevaan kuulojärjestelmään. Nämä ytimet muodostavat myös sisäkorvaa hermottavan olivokokleaarisen nipun lähteen. Marsulla nousevat projektiot colliculi inferioriin ovat pääasiassa ipsilateraalisia (>80 %), ja suurin yksittäinen lähde tulee SPON:sta. Myös ventraaliset ytimet (RPO, VMPO, AVPO, & VNTB) ovat lähes kokonaan ipsilateraalisia, kun taas loput ytimet projisoivat bilateraalisesti.
Ventral Nucleus of Trapezoid Body (VNTB)
- VNTB on pieni ydin, joka sijaitsee lateraalisesti MNTB:hen nähden, ja ventraalisesti MSO:n vieressä.
- Tämä heterogeenisesta solupopulaatiosta koostuva ydin heijastuu moniin kuuloytimiin ja muodostaa mediaalisen olivokokleaarisen nipun (MOC), joka innervoi sisäkorvan ulompia karvasoluja . Nämä solut sisältävät elektromotilisia kuituja, ja ne toimivat mekaanisina vahvistimina/vaimentimina sisäkorvassa.
- Tydin projisoituu molempiin IC:iin, eikä yksikään solu projisoi molemminpuolisesti.
Trapetsirungon lateraalinen ydin (LNTB, Lateral Nucleus of the Trapezoid Body)
- Sijaitsee ventriaalisesti LSO:n kohdalla
- AVCN:n pallomaiset puskasolut projisoivat kollateraalit bilateraalisesti ja pallomaiset puskasolut projisoivat kollateraalit ipsilateraalisesti LNTB:n neuroneihin.
- Solut ovat immunoreaktiivisia glysiinille, ja ne leimataan retrogradisesti tritioidun glysiinin injektion jälkeen LSO:han
- Tydin projisoituu molempiin IC:iin, ja vain harvat solut projisoituvat bilateraalisesti, sekä ipsilateraaliseen LSO:han.
- Suuret monipolaariset solut projisoituvat sekä kissan että marsun sisäkorvaperäiselle ytimelle mutta eivät IC:lle.
- Sisääntulot tapahtuvat usein pidettyjen päätyjen kautta, mikä tuottaa hyvin nopean signaalinsiirron.
Superior Periolivary Nucleus (SPON) (Dorsomedial Periolivary Nucleus (DMPO))
- Sijoittuu suoraan dorsaalisesti MNTB:hen
- Rotilla SPON on homogeeninen GABAerginen ydin. Nämä tonotopisesti järjestäytyneet neuronit saavat eksitatorisia sisääntuloja kontralateraalisen ventraalisen sisäkorvan ytimen oktopus- ja multipolarisoluista soluista, glyserergisen (inhibitorisen) sisääntulon ipsilateraalisesta MNTB:stä, tuntemattoman GABAergisen (inhibitorisen) sisääntulon ja projisoituvat ipsilateraaliseen ICC:hen. Useimmat neuronit reagoivat vain ärsykkeen offset-hetkellä, pystyvät lukkiutumaan AM-ärsykkeisiin jopa 200 Hz:iin asti, ja ne voivat muodostaa perustan ICC:n kestoselektiivisyydelle. Huomionarvoista on, että SPON-neuronit eivät saa laskevia syötteitä IC:stä, eikä se projisoi sisäkorvaan tai sisäkorvan ytimeen, kuten monet periolivaryhmän ytimet tekevät.
- Marsussa pyöreät tai soikeat monipolaariset solut projisoituvat molempiin IC:iin, ja monet solut projisoituvat bilateraalisesti. Pidemmät solut, jotka projisoivat sisäkorvan ytimeen, eivät projisoi ICC:hen. Näyttää siltä, että on olemassa kaksi solupopulaatiota, joista toinen projisoituu ipsilateraalisesti ja toinen bilateraalisesti.
- Suurin osa tiedoista oli peräisin jyrsijöiden SPON:sta, johtuen ytimen huomattavasta koosta näissä lajeissa, ja kissan DMPO:lla on tehty hyvin vähän tutkimuksia, joista yksikään ei ollut laaja.
Dorsal Periolivary Nucleus (DPO)
- Sijoittuu dorsaalisesti ja mediaalisesti LSO:han nähden
- Sisältää sekä EE- (molempien korvien herättämiä) että E0-yksiköitä (vain kontralateraalisen korvan herättämiä).
- Neuronit ovat tonotopisesti järjestäytyneitä ja korkeataajuisia.
- Voi kuulua yhteen ytimeen yhdessä DLPO:n kanssa
- Tydin projisoituu molempiin IC:iin, eikä yhtään solua projisoi bilateraalisesti.
Dorsolateraalinen periolivulaarinen ydin (DLPO)
- Sijoittuu dorsaalisesti ja lateraalisesti LSO:sta
- Sisältää sekä EE- (molempien korvien herättämiä) että E0-yksiköitä (vain kontralateraalisen korvan herättämiä).
- Neuronit ovat tonotopisesti järjestäytyneitä ja matalataajuisia.
- Voi kuulua yhteen ytimeen yhdessä DPO:n kanssa
- Tydin projisoituu molempiin IC:iin, ja harvat solut projisoituvat bilateraalisesti.
Ventrolateraalinen periolivaarinen ydin (VLPO)
- Sijoittuu ventral ja LSO:n ventraalisen kukkulan sisällä
- Sisältää sekä EI- (kontralateraalisen korvan herättämiä ja ipsilateraalisen korvan estämiä) että E0-yksikköjä (herättämiä vain kontralateraalisesta korvasta).
- Neuronit ovat tonotopisesti järjestäytyneitä ja korkeataajuisia.
Anterolateraalinen periolivaarinen ydin (ALPO)
- Tydin projisoituu molempiin IC:iin, eikä yhtään solua projisoi bilateraalisesti.
- Suuret monipolaariset solut projisoituvat sekä kissalla että marsulla sisäkorvaperäiseen ytimeen, mutta eivät IC:hen.
Ventromediaalinen periolivarinen ydin (VMPO)
- Sijaitsee MSO:n ja MNTB:n välissä.
- Lähettää projektioita ICC:hen bilateraalisesti.
- Tydin projisoi molempiin IC:iin, eikä yksikään solu projisoi bilateraalisesti.
Rostral periolivary nucleus (RPO) (Anterior periolivary nucleus (APO))
- Sijaitsee MSO:n rostraalisen napan ja VNLL:n välissä
Caudal Periolivary Nucleus (CPO) (Posterior Periolivary Nucleus (PPO))
- Sijaitsee MSO:n kaudaalisen napan ja kasvojen ytimen (7N) välissä
Posteroventraalinen periolivaarinen ydin (PVPO)
- Tydin projisoituu molempiin IC:iin, eikä yhtään solua projisoi bilateraalisesti.
Patofysiologia
Autismia, epilepsiaa ja kehitysvammaisuutta sairastaneen 21-vuotiaan naisen ruumiinavauksessa todettiin lähes täydellinen oliivin yläosan puuttuminen.
Katso myös
- Olivary body
- Kulesza RJ, Cytoarchitecture of the human superior olivary complex: Medial and lateral superior olive. Hearing Research 225(2007) 80-90
- 2.0 2.1 Oliver DL, et al. Axonal projections from the lateral and medial superior olive to the inferior colliculus of the cat: a study using electron microscopic autoradiography. J Comp Neurol. 1995 Sep 11;360(1):17-32 Cite error:
- Cajal, S. R. Y. ja L. Azoulay (1909). Histologie du système nerveux de l’homme et des vertébrés. Paris, Maloine.
- 4.0 4.1 Kandel, et al Principles of Neuroscience. Neljäs painos. s. 591-624. Copyright 2000, by McGraw-Hill Co. Cite error: Invalid
<ref>tag; name ”R2” defined multiple times with different content - Kulesza, R. J., Jr. (2007). ”Cytoarchitecture of the human superior olivary complex: medial and lateral superior olive”. Hear Res 225(1-2): 80-90.
- Tsuchitani, C. ja J. C. Boudreau (1967). ”Ärsykkeen taajuuden ja voimakkuuden koodaus kissan ylemmän oliivin S-segmentin soluissa”. J Acoust Soc Am 42(4): 794-805.
- 8.0 8.1 8.2 Cite error: Invalid
<ref>tag; name ”R10” defined multiple times with different content - Wu, S. H. and J. B. Kelly (1994). ”Fysiologiset todisteet ipsilateraalisesta inhibitiosta lateraalisessa ylemmässä oliivissa: synaptiset vasteet hiiren aivoleikkeessä”. Hear Res 73(1): 57-64.
- Brownell, W. E., P. B. Manis, et al. (1979). ”Ipsilateraaliset estävät vasteet kissan lateraalisessa ylemmässä oliivissa.” Brain Res 177(1): 189-93.
- Darrow, K. N., S. F. Maison, et al. (2006). ”Sisäkorvan efferentti palaute tasapainottaa interauraalista herkkyyttä.” Nat Neurosci 9(12): 1474-6.
- Warr, W. B. ja J. J. Guinan, Jr. (1979). ”Corti-elimen efferentti innervaatio: kaksi erillistä järjestelmää”. Brain Res 173(1): 152-5.
- 13.0 13.1 Schofield, B. R. ja N. B. Cant (1991). ”Superior olivary complexin organisaatio marsulla. I. Sytoarkkitehtuuri, sytokromioksidaasin histokemia ja dendriittinen morfologia”. J Comp Neurol 314(4): 645-70. Cite error: Invalid
<ref>tag; name ”R7” defined multiple times with different content - 14.0 14.1 14.2 14.3 14.4 14.5 14.6 14.7 14.8 Illing, R. B., K. S. Kraus, et al. (2000). ”Superior olivary complexin plastisuus”. Microsc Res Tech 51(4): 364-81. Cite error: Cite error: Invalid
<ref>tag; name ”R5” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R5” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R5” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R5” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R5” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R5” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R5” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R5” defined multiple times with different content - Warr, W. B. and J. E. Beck (1996). ”Moninkertaiset projektiot trapetsirungon ventraalisesta ytimestä rotalla”. Hear Res 93(1-2): 83-101.
- 16.00 16.01 16.02 16.03 16.04 16.05 16.06 16.07 16.08 16.09 16.10 Schofield, B. R. ja N. B. Cant (1992). ”Superior olivary complexin organisaatio marsulla: II. Projektiokuviot periolivaryhmän ytimistä colliculus inferioriin”. J Comp Neurol 317(4): 438-55. Cite error: Cite error: Invalid
<ref>tag; name ”R6” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R6” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R6” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R6” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R6” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R6” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R6” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R6” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R6” defined multiple times with different content Cite error: Invalid<ref>tag; name ”R6” defined multiple times with different content - Smith, P. H., P. X. Joris, et al. (1993). ”Fysiologisesti karakterisoitujen pallomaisten puskurisolujen aksonien projisoinnit kissan sisäkorvan ytimestä: todisteita viivelinjoista mediaaliseen ylempään oliiviin.” J Comp Neurol 331(2): 245-60.
- Wenthold, R. J., D. Huie, et al. (1987). ”Glysiini-immunoreaktiivisuus lokalisoituu sisäkorvan ytimeen ja ylempään oliivikompleksiin.” Neuroscience 22(3): 897-912.
- 19.0 19.1 Adams, J. C. (1983). ”Periolivariaalisolujen sytologia ja niiden projektioiden järjestäytyminen kissalla.” J Comp Neurol 215(3): 275-89. Cite error: Invalid
<ref>tag; name ”R8” defined multiple times with different content - Friauf, E. and J. Ostwald (1988). ”Fysiologisesti luonnehdittujen rotan ventraalisen sisäkorvan ytimen neuronien divergentit projektiot, jotka osoitetaan piparjuuriperoksidaasin aksoninsisäisellä injektiolla”. Exp Brain Res 73(2): 263-84.
- Kulesza, R. J., Jr. ja A. S. Berrebi (2000). ”Rotan ylempi paraolivaarinen ydin on GABAerginen ydin”. J Assoc Res Otolaryngol 1(4): 255-69.
- Kulesza, R. J., Jr., G. A. Spirou, et al. (2003). ”Neuronien fysiologiset vasteominaisuudet rotan ylemmässä paraolivardisessa ytimessä”. J Neurophysiol 89(4): 2299-312.
- WHITE JS, WARR WB. Olivokokleaarisen nipun kaksoisalkuperä albiino-rotalla. J. Comp. Neurol. 219:203-214, 1983.
- Guinan, J. J., Jr., S. S. Guinan, et al. (1972). ”Yksittäiset kuuloyksiköt ylemmässä olivarykompleksissa. I. Vastaukset ääniin ja fysiologisiin ominaisuuksiin perustuvat luokittelut”. Int J Neurosci 4: 101-20.
- Tsuchitani, C. (1977). ”Kissan superior olivary complexin lateraalisten soluryhmien toiminnallinen järjestäytyminen”. J Neurophysiol 40(2): 296-318.
- Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996). Autismin embryologinen alkuperä: aivohermojen motoristen ytimien kehityspoikkeavuudet. J Comp Neurol 370 (2): 247-61.
- MedEd at Loyola Neuro/frames/nlBSs/nl27fr.htm
- Image at med.utah.edu (pick 9b. Pons)
Tämä artikkeli perustuu alunperin Gray’s Anatomy -teoksen julkiseen painokseen. Näin ollen osa sen sisältämistä tiedoista saattaa olla vanhentuneita. Muokkaa artikkelia, jos näin on, ja poista tämä ilmoitus, kun se ei ole enää merkityksellinen.
|
v-d-e
Aivot: Rhombencephalon (takaraivot) |
||||||
|---|---|---|---|---|---|---|
| Metencephalon/ pons |
|
Dorsaalinen/
(tegmentum)
|
Ventraalinen/
(tyvi) |
Raphe/retikulaarinen
|
||
| Myelencephalon/ medulla |
|
Dorsaalinen
pinta: Posterior median sulcus – Postero-lateraalinen sulcus – Area postrema kallon ytimet: GVA: VII,IX,X: Solitary/tractus – SVA: Gustatory nucleus – GSE: XII – GVE: IX,X,XI: Ambiguus – SVE: X: Dorsaalinen – IX: Inferiorinen syljeneritysydin – MLF, III, IV ja VI sensorinen/ascending: Gracile nucleus – Cuneate nucleus (Accessory cuneate nucleus) – Sensory decussation – Medial lemniscus motorinen/laskeva: Dorsaalinen hengitysryhmä
|
Ventraalinen
|
Raphe/retikulaarinen
|
||
|
v-d-e
Aistijärjestelmä: Kuulo- ja tasapainojärjestelmä (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ulkokorva |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Korvalehti) – Korvakäytävä – Korvalihakset Korvalehti (Umbo, Pars flaccida) |
|||||||||||
| Keskikorva |
|
|||||||||||
| Sisäkorva/ (kalvomainen labyrintti, luinen labyrintti) |
|
|||||||||||
| {| class=”navbox collapsible nowraplinks” style=”margin:auto;” | ||||||||||||
|
—
|
||||||||||||
|
||||||||||||
|}
Tällä sivulla käytetään Wikipedian Creative Commons -lisensoitua sisältöä (Näytä tekijät).
Vastaa