Shark Week: Mitä Megalodon söi?
On 13 joulukuun, 2021 by admin Vieraileva kirjoittaja: Jack Cooper, MSc
UoB Graduate / PhD Student, University of Swansea
Kuten olemme tähän mennessä nähneet, Megalodonin taksonomista luokittelua ja maksimikokoa koskevaan tieteelliseen yksimielisyyteen on tullut muutoksia ja päivityksiä. Sen lempivälipalasta ollaan kuitenkin tiedeyhteisössä yleisesti yhtä mieltä. Toinen harvinaisuus on se, että se on asia, joka on yleensä kuvattu melko tarkasti fiktiossa. Sallikaa minun esitellä hai, joka söi valaita – oikeita valaita!
Monet nykyisistä suurista makrosyöjähaista tunnetaan siitä, että ne syövät merinisäkkäitä, erityisesti vanha ystävämme valkohai. Joissakin ikonisimmissa valokuvissa tämä hai heittäytyy vedestä viimeisin uhri leukojensa välissä (kuva 1). Nuoret valkohait eivät kuitenkaan syö hylkeitä heti alusta alkaen. Sen sijaan ne alkavat syödä kaloja ja pienempiä haita. Jos tarkastelemme jälleen Gatunin muodostumaa Panamassa, löydämme runsaasti fossiilisia haita, rauskuja ja otoliitteja (kalojen sisäkorvista peräisin olevia rakenteita) sekä runsaasti selkärangattomia eläimiä, kuten nilviäisiä .
Mitä emme kuitenkaan löydä, ovat fossiiliset merinisäkkäät. Tämä näyttäisi viittaavan siihen, että Megalodonin poikaset söivät myös kaloja ja pienempiä haita sen sijaan, että olisivat heti ryhtyneet jahtaamaan merinisäkkäitä. Tätä kutsutaan ontogeneettiseksi siirtymäksi ruokavaliossa, mikä tarkoittaa sitä, että hai vaihtaa suosimaansa ravinnonlähdettä kasvaessaan. Syömällä suurempaa ruokaa ja todennäköisesti paljon sitä, kun Megalodon saavutti aikuiskokonsa, se olisi pystynyt pitämään yllä niin suuren kokonsa aiheuttamia aineenvaihdunnallisia vaatimuksia. Se olisi voinut tehdä tämän syömällä valaiden lisäksi lähes mitä tahansa, mihin se olisi törmännyt.

Kun otetaan huomioon, miten paljon ruokaa Megalodonin olisi pitänyt syödä, jotta se olisi voinut perustella valtavan kokonsa, se oli lähes varmasti opportunistinen syöjä. Fossiiliaineisto on paras oppaamme tässä asiassa. Tämä johtuu jälkifossiileista, jotka ovat geologisia merkintöjä biologisesta toiminnasta. Ne voivat vaihdella dinosaurusten jalanjäljistä luissa oleviin naarmuihin, jotka osoittavat, että jokin puri eläintä, jolle luu kuului. Ja Megalodon jätti jälkeensä muutamia tällaisia jälkifossiileja, jotka tarjoavat keskeisiä todisteita sen tappamisista.
Monista eri valaille kuuluvista fossiileista on löydetty huomattavia puremajälkiä. Yksi hyvin siteerattu esimerkki on Marylandissa sijaitsevasta Yorktownin muodostumasta, jossa useissa valaan luissa oli puremajälkiä, jotka oli selvästi tehty suurilla hammastetuilla hampailla . Itse asiassa Megalodonin hampaita on joskus löydetty suoraan näiden luiden yhteydessä.
Hait menettävät ja vaihtavat hampaita tuhansia kertoja elämänsä aikana, ja siksi hait menettävät usein hampaita tehdessään väkivaltaisen tapon. Tämä on keskeinen syy siihen, miksi hain hampaat ovat niin yleisiä fossiileja. Hain hampaiden esiintyminen aivan naarmuuntuneen valaan luun vieressä viittaa kyllä siihen, että ruokailu oli käynnissä. Paleontologien olisi kuitenkin aina pohdittava, kun tällaisia fossiileja löydetään: oliko kyseessä todella saalistus? Vai oliko kyseessä kenties haaskaaminen?
Haaskaaminen on yleinen ravintokäyttäytyminen nykyaikaisilla makrosyöjähaiden, kuten valkohaiden (kuva 2) ja tiikerihaiden, kaltaisilla haikaloilla. Kun nämä hait törmäävät pinnalla kelluviin kuolleisiin valaisiin, se on all-you-can-eat buffet. Kriittistä on, että haiden syöminen ei vaadi energiankulutusta, joka voi mennä hukkaan epäonnistuneessa saalistusyrityksessä. Kun otetaan huomioon, että Megalodonin oli pystyttävä säilyttämään valtava kokonsa, ei olisi kovin järkevää jättää ilmaista ateriaa väliin, jos se kohtasi kuolleen valaan. Näin ollen sen perusteella, miten suuret nykyaikaiset hait syövät, useimmat meistä olisivat valmiita lyömään vetoa, että Megalodon luultavasti söi raatoja, kun siihen tarjoutui tilaisuus.

Tästä huolimatta on olemassa yksi erityisen merkittävä fossiili, joka viittaa Megalodonin saalistusyritykseen. Ja ratkaisevaa on, että tiedämme tämän ennen kaikkea siksi, että sen yritys epäonnistui. Vuonna 2010 julkaistussa artikkelissa kuvattiin Pohjois-Carolinassa sijaitsevasta Yorktownin muodostumasta (jep, kahdesta Yorktownin muodostumasta) peräisin oleva osittainen plioseenisen kylkiluun fossiili, joka kuului valaaseen. Myös tässä fossiilissa oli puremajälkiä muistuttavia vaurioita, mutta erityisesti näiden jälkien päällä näytti olevan kudottu luupeite. Kirjoittajat tulkitsivat tämän merkiksi infektiosta, joka oli syntynyt eläimen toipuessa puremajälkien aiheuttamasta luuvammasta. Nämä jäljet sopivat hyvin yhteen sahalaitaisen pureman kanssa, mikä viittaa siihen, että hyökkääjällä oli sahalaitaiset hampaat – aivan kuten Megalodonilla. Tutkimus siis viittasi siihen, että Megalodon tai jokin muu suuri hai oli hyökännyt eläimen kimppuun, josta fossiili oli peräisin, mutta uhri oli selvinnyt yrityksestä vain kuollakseen kuuden viikon kuluttua vamman aiheuttamaan infektioon.
Aivan hiljattain Perun Pisco-muodostelmasta löydettiin ensimmäiset Megalodoniin liittyvät fossiiliset puremajäljet eteläiseltä pallonpuoliskolta . Ehkä se oli tarkoituskin, sillä tämä on sama paikka, josta Gordon Hubbell löysi poikkeuksellisen Carcharodon-fossiilin (jota käsitellään Q1:ssä). Täältä löydetty purema-aineisto sisälsi kallon ja kylkiluiden jäänteitä, mikä viittaa siihen, että hai oli purrut kohdettaan sivusta. Merkittävintä on, että jäänteet tunnistettiin ensimmäistä kertaa yksittäiseksi lajiksi – laji oli Piscobalaena nana, pieni paaluvala. Kuten muissakin jälkifossiileissa, puremajäljet sopivat hyvin yhteen suurten sahalaitaisten hampaiden kanssa – hyvä peruste syyttää Megalodonia hyökkäyksestä (kuva 3). Näytti siis siltä, että Megalodon pystyi kohdistamaan hyökkäyksensä myös pienempiin valaisiin (oli kyse sitten saalistuksesta tai haaskauksesta), mikä vastaa hyvin pitkälti ehdotuksia, joiden mukaan se todennäköisesti kohdistui myös delfiineihin, merikilpikonniin ja muihin pienempiin saaliseläimiin .

Vaikka saalistusstrategioita on vaikea tulkita fossiileista, nykyaikaiset hait voivat antaa meille käsityksen siitä, miten Megalodon metsästi. Sen puremavoima olisi ollut poikkeuksellisen voimakas vaurioittamaan valaan luita. Valkohain kallon tietokonemallissa on ehdotettu, että sen purentavoima ylittää noin 1,8 tonnia, mikä tekee siitä yhden kaikkien elävien eläinten voimakkaimmista purentavoimista. Tämän tutkimuksen mukaan Megalodonin puremavoima oli vielä suurempi, arviolta 108 514-182 201 N.
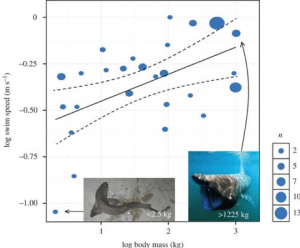
Tutkijat ovat myös yrittäneet laskea Megalodonin uintinopeutta, mikä voi auttaa saamaan tietoa siitä, miten se metsästi. Hailla voi olla purskahdusnopeus, joka on nopean uinnin nopea sysäys, jota käytetään tyypillisesti saaliin nappaamiseen. Niiden tavanomainen nopeus satunnaisessa uinnissa on kuitenkin se, mitä kutsumme jatkuvaksi uintinopeudeksi. Tämä vaihtelee lajeittain, mutta eräässä David Jacobyn johtamassa tutkimuksessa yhdistettiin 64 aiemmasta tutkimuksesta saatuja tietoja 26 lajista ja kehitettiin malli, joka ennustaa jatkuvan uintinopeuden ja hain aineenvaihdunnan ja ruumiinmassan välisen skaalautuvan suhteen. Heidän mallinsa, johon oli sisällytetty aineenvaihdunta, osoitti skaalautuvaksi eksponentiksi 0,173, joka on tiivistettynä seuraava yhtälö:
Uintinopeus (ms ¯¹) ∝ Massa (kg)^0,173
Kun he kuitenkin korjasivat tiedot fylogeneesin huomioon ottamiseksi, he havaitsivat, että minimiuintinopeudet skaalautuivat ruumiinmassan kanssa hieman alhaisemmalla eksponentilla, joka oli 0,15. Yllä olevassa yhtälössä oletetaan lisäksi, että vakio on 1, jotta se vastaisi mittasuhteita, mutta aineisto paljasti, että tämä vakio oli itse asiassa 0,266. Sen vuoksi he konstruoivat tehoyhtälön:
Uintinopeus (ms ¯¹) = 0,266 Massa (kg) ^0,15
Tästä he pystyvät laskemaan Megalodonin jatkuvan uintinopeuden sen massasta riippuen. Esimerkiksi Gottfriedin työssä 15,9 metrin pituisen hain massaksi laskettu ~48 000 kg painava Megalodon paljastaisi hain uivan jatkuvasti 1,34 m/s – noin 4,8 km/h . Soveltamalla tätä 52 000 kg painavaan Megalodoniin saadaan jatkuvaksi uintinopeudeksi 1,36 m/s eli 4,9 km tunnissa, kun taas jos mentäisiin todella äärimmäisyyksiin ja simuloitaisiin 100 000 kg painavaa haita, tämän mallin mukaan uintinopeus olisi 1,50 m/s eli 5,4 km tunnissa. Tämä malli on osoittautunut erittäin tehokkaaksi paitsi Megalodonin uintinopeuden laskemisessa myös mallin tarjoamisessa useille hailajeille (kuva 4) .
Hait käyttävät kuitenkin juuri purskahdusnopeutta napatakseen nopeasti saaliinsa. Valkohait hyödyntävät tätä ”breaching-käyttäytymisessään”, jossa ne ponnistavat vedestä syöksyen saalista alhaalta yllättääkseen sen (kuva 1) . Ne pystyvät tähän, koska ne ovat hyvin naamioituneita vastavärjäyksen ansiosta. Koska Megalodon oli todennäköisesti myös vastavärjätty ja sillä oli samanlainen ekologia ja ruumiinrakenne kuin valkohailla , herää kysymys: kykenikö Megalodon myös breikkaamaan?
Mahdollisesti. Valkohain puhkeamisnopeus perustuu fysiologiseen sopeutumiseen nimeltä mesotermia . Tämä on jotain, mitä Megalodonilla on hiljattain ehdotettu olleen myös (käsitellään yksityiskohtaisesti Q5:ssä). Mesotermisillä kaloilla on yleensä nopeampi uintinopeus kuin ektotermisillä kaloilla , ja tämän perusteella on ehdotettu, että Megalodonilla saattoi olla jopa 37,15 km/h nopeus.
Tämä uskomattoman nopea nopeus näin suurelle haille saattoi hyvinkin riittää, jotta se olisi pystynyt murtautumaan. Tämän lisäksi myös ruskohain (Cetorhinus maximus), nykyisin toiseksi suurimman hain, on havaittu breikkaavan . Ehkä jättiläismegalodon pystyi todellakin hyppäämään vedestä, kun se nappasi viimeisimmän ateriansa. Emme vain tiedä sitä vielä, mutta henkilökohtainen ja (jokseenkin) ammattimainen mielipiteeni on, että se saattoi hyvinkin olla mahdollista.
Mikäli Megalodonilla oli näin laaja ruokalista, Megalodon oli huippupetohai, jolla ei ollut omia tunnettuja saalistajia. Tämä tarkoittaa kahta tärkeää asiaa. Ensinnäkin sillä oli todennäköisesti kova kilpailu saaliistaan, mikä saattoi lopulta vaikuttaa sen sukupuuttoon kuolemiseen. Toiseksi huippupetona Megalodon olisi ollut äärimmäisen tärkeä ekosysteemilleen, sillä se olisi pitänyt saaliskantansa kurissa ja siten säilyttänyt terveen ekosysteemin.
Sen vuoksi nykypäivän hait ovat niin tärkeitä meriympäristöille. Huippupetohain häviäminen aiheuttaa peruuttamattomia kaskadivaikutuksia, jotka muuttavat sen jälkeensä jättämää yhteisöä . Näin ollen meriekosysteemimme ovat sellaisia kuin ne ovat osittain siksi, että Megalodon ei ole enää osa niitä. Ja nykyisten valtamerten valtavat valaat voivat sen seurauksena levätä rauhassa.
- Estrada JA, Rice AN, Natanson LJ & Skomal GB 2006. Nikamien isotooppianalyysin käyttö valkohaiden ontogeneettisen ravinto-ekologian rekonstruoinnissa. Ecology 87, 829-834.
- Pimiento C, Ehret DJ, MacFadden BJ & Hubbell G 2010. Sammuneen jättiläishain Megalodonin muinainen kasvualue Panaman mioseenistä. PLoS One 5, e10552.
- Pimiento C, González-Barba G, Ehret DJ, Hendy AJ, MacFadden BJ & Jaramillo C 2013. Hait ja rauskut (Chondrichthyes, Elasmobranchii) Panaman myöhäismiocene Gatunin muodostumasta. J. Paleontol. 87, 755-774.
- Purdy RW 1996. Fossiilisten valkohaiden paleoekologia. In: Great White Sharks: The Biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 67-78, Academic Press, San Diego.
- Aguilera OA, García L & Cozzuol MA 2008. Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation. Paläontol. Z. 82, 204-208.
- Fallows C, Gallagher A. & Hammerschlag N 2013. Valkoisten haiden (Carcharodon carcharias) valaanpyynti ja sen mahdollinen rooli huippupedon ekologian muokkaamisessa edelleen. PLoS One, 8, e60797.
- Dudley SFJ, Anderson-Reade MD, Thompson GS & McMullen PB 2000. Valashaiden, Carcharodon carcharias, ja tiikerihaiden, Galeocerdo cuvier, samanaikainen valaan raadon syönti. Fish. Bull. 98, 646-649.
- Kallal RJ, Godfrey SJ & Ortner DJ 2010. Plioseenisen valaan kylkiluun luureaktiot osoittavat lyhytaikaista selviytymistä saalistustapahtumasta. Int. J. Osteoarchaeol. 22, 253-260.
- Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M & Bianucci G 2017. Tähtäsikö sukupuuttoon kuollut jättiläishai Carcharocles megalodon pieniin saaliisiin? Puremajäljet merinisäkkäiden jäänteissä Perun myöhäismioseenistä. Palaeogeogr. Palaeoclimatol. Palaeoecol. 469, 84-91.
- Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, Ferrara TL, Cunningham E, Dean MN & Summers AP 2008. Kolmiulotteinen tietokoneanalyysi valkohain leukamekaniikasta: kuinka kovaa valkohai voi purra? J. Zool. 276, 336-342.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2015. Onko haiden uintinopeuden skaalautuminen aineenvaihdunnan ohjaamaa? Biol. Lett. 11, 20150781.
- Gottfried MD, Compagno LJV & Bowman SC 1996. Jättimäisen ”megahampaan” hain Carcharodon megalodonin koko ja luuston anatomia. In: Great White Sharks: The Biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 55-66, Academic Press, San Diego.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2016. Korjaus artikkeliin ”Is the scaling of swim speed in sharks driven by metabolism?”. Biol. Lett. 12, 20160775.
- Watanabe YY, Payne NL, Semmens JM, Fox A & Huveneers C 2019. Endotermisten valkohaiden uintistrategiat ja energetiikka ravinnonhankinnan aikana. J. Exp. Biol. 222, jeb185603.
- Cooper JA, Pimiento C, Ferrón HG & Benton MJ (in press). Sammuneen jättiläishain Otodus megalodonin ruumiin mitat: A 2D reconstruction. Sci. Rep.
- Watanabe YY, Goldman KJ, Caselle JE, Chapman DD & Papastamatiou YP 2015. Vertailevat analyysit eläinten seuranta-aineistosta paljastavat kalojen endotermian ekologisen merkityksen. Proc. Nat. Acad. Sci. 112, 6104-6109.
- Ferrón HG 2017. Alueellinen endotermia jättiläismäisyyden laukaisijana joillakin sukupuuttoon kuolleilla makropredatorisilla haikaloilla. PLoS One, 12, e0185185.
- Johnston EM, Halsey LG, Payne NL, Kock AA, Iosilevskii G, Whelan B & Houghton JD 2018. Poikkeuksellisten breikkaustapahtumien paljastama piilevä voima paistattelevissa haikaloissa (Latent power of basking sharks revealed by exceptional breaching events). Biol. Lett. 14, 20180537.
- Myers RA, Baum JK, Shepherd TD, Powers SP & Peterson CH 2007. Kaskadivaikutukset, jotka aiheutuvat huippusaalistavien haiden häviämisestä rannikkomerestä. Science, 315, 1846-1850.
Edited by Rhys Charles
Vastaa