N-sidoksinen glykosylaatio
On 18 joulukuun, 2021 by admin
N-sidonnaisten glykaanien biosynteesi tapahtuu kolmen päävaiheen kautta:
- Doliin sidoksissa olevan esiaste-oligosakkaridin synteesi
- En bloc-siirto esiaste-oligosakkaridista proteiiniin
- Oligosakkaridin prosessointi
Synteesi, en bloc-siirto ja esiaste-oligosakkaridin alustava trimmaus tapahtuvat endoplasmisessa retikulumissa (ER). Oligosakkaridiketjun myöhempi prosessointi ja modifiointi tapahtuu Golgi-laitteistossa.
Glykoproteiinien synteesi on siten alueellisesti erotettu eri solukompartimentteihin. Näin ollen syntetisoituvan N-glykaanin tyyppi riippuu sen saatavuudesta näissä solukompartimenteissa oleville eri entsyymeille.
Vaihtelevuudesta huolimatta kaikki N-glykaanit syntetisoituvat yhteisen reitin kautta, ja niillä on yhteinen ydinglykaanirakenne.
Ydinglykaanirakenne koostuu pääosin kahdesta N-asetyyliglukosamiini- ja kolmesta mannoosijäännöksestä. Tätä ydinglykaania työstetään ja muokataan edelleen, jolloin syntyy monenlaisia N-glykaanirakenteita.
Prekursorioligosakkaridin synteesiEdit
N-sidoksisen glykosylaation prosessi alkaa doliiniin sidotun GlcNAc-sokerin muodostumisella. Doliholi on lipidimolekyyli, joka koostuu toistuvista isopreeniyksiköistä. Tämä molekyyli löytyy kiinnittyneenä ER:n kalvoon. Sokerimolekyylit kiinnittyvät doliholiin pyrofosfaattisidoksella (yksi fosfaatti on alun perin sitoutunut doliholiin, ja toinen fosfaatti on peräisin nukleotidisokerista). Tämän jälkeen oligosakkaridiketjua jatketaan lisäämällä eri sokerimolekyylejä vaiheittain, jolloin muodostuu esiasteoligosakkaridi.
Tämän esiasteoligosakkaridin kokoaminen tapahtuu kahdessa vaiheessa: Vaihe I ja II. Vaihe I tapahtuu ER:n sytoplasmisella puolella ja vaihe II tapahtuu ER:n luminaalisella puolella.
Prekursorimolekyyli, joka on valmis siirrettäväksi proteiiniin, koostuu 2 GlcNAc-, 9 mannoosi- ja 3 glukoosimolekyylistä.

|
|
| Vaihe II | |
on mannoosijäämien luovuttaja (muodostuminen : Dol-P + GDP-Man → Dol-P-Man + GDP) ja Dol-P-Gluc on glukoosijäämien luovuttaja (muodostuminen : Dol-P + UDP-Glc → Dol-P-Glc + UDP).
|
|
Glykaanin siirtyminen proteiiniinEdit
Kun esiaste-oligosakkaridi on muodostunut, valmis glykaani siirretään sitten nasentoituvaan polypeptidiin ER:n kalvon lumenissa. Tätä reaktiota ohjaa energia, joka vapautuu doligoli-glykaanimolekyylin välisen pyrofosfaattisidoksen pilkkoutumisesta.On kolme ehtoa, joiden on täytyttävä, ennen kuin glykaani siirretään nasentoituvaan polypeptidiin:
- Asparagiinin on sijaittava primäärirakenteessa tietyssä konsensussekvenssissä (Asn-X-Ser tai Asn-X-Thr tai harvinaisissa tapauksissa Asn-X-Cys).
- Asparagiinin on sijaittava sopivasti proteiinin kolmiulotteisessa rakenteessa (sokerit ovat polaarisia molekyylejä, joten niiden on oltava kiinnittyneinä asparagiiniin, joka sijaitsee proteiinin pinnalla eikä hautautuneena proteiinin sisälle)
- Asparagiinin on sijaittava endoplasmisen retikulumin luminaalisella puolella, jotta N-sidoksinen glykosylaatio voi alkaa. Kohdejäännökset löytyvät joko erittyvistä proteiineista tai transmembraaniproteiinin alueilta, jotka ovat lumeniin päin.
Oligosakkaryylitransferaasi on entsyymi, joka on vastuussa konsensussekvenssin tunnistamisesta ja esiaste-glykaanin siirtämisestä polypeptidien akseptoriin, joka translatioidaan endoplasmisen retikulumin lumenissa. Prekursorimolekyylin alustava trimmaus tapahtuu ER:ssä ja myöhempi prosessointi tapahtuu Golgissa.
Valmiin glykaanin siirtyessä nasentoituvaan polypeptidiin rakenteesta poistetaan kaksi glukoosijäännöstä. Glykosidaaseiksi kutsutut entsyymit poistavat joitakin sokerijäännöksiä. Nämä entsyymit voivat rikkoa glykosidisidoksia vesimolekyylin avulla. Nämä entsyymit ovat eksoglykosidaaseja, koska ne toimivat vain glykaanin pelkistämättömässä päässä sijaitseviin monosakkaridijäämiin. Tämän alkuperäisen trimmausvaiheen ajatellaan toimivan laadunvalvontavaiheena ER:ssä proteiinin taittumisen valvomiseksi.
Kun proteiini on taittunut oikein, glukosidaasi I ja II poistavat kaksi glukoosijäännöstä. Viimeisen kolmannen glukoosijäännöksen poistaminen ilmoittaa, että glykoproteiini on valmis siirtymään ER:stä cis-Golgiin. ER-mannosidaasi katalysoi tämän viimeisen glukoosin poiston. Jos proteiini ei kuitenkaan taitu oikein, glukoosijäämiä ei poisteta, eikä glykoproteiini näin ollen voi poistua endoplasmisesta retikulumista. Chaperoniproteiini (kalnexiini/kalretikuliini) sitoutuu taittumattomaan tai osittain taittuneeseen proteiiniin auttaakseen proteiinin taittumista.
Seuraavaan vaiheeseen kuuluu sokerijäämien lisäys ja poisto cis-Golgissa. Näitä modifikaatioita katalysoivat vastaavasti glykosyylitransferaasit ja glykosidaasit. Cis-Golgissa joukko mannosidaaseja poistaa osan tai kaikki neljästä mannoosijäännöksestä α-1,2-sidoksissa. Kun taas Golgin mediaalisessa osassa glykosyylitransferaasit lisäävät glykaanin ydinrakenteeseen sokerijäännöksiä, jolloin syntyy kolmenlaisia glykaaneja: runsaasti mannoosia sisältäviä, hybridi- ja kompleksisia glykaaneja.
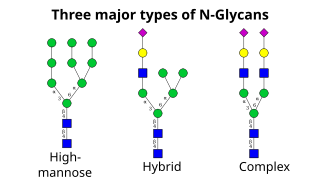
- Korkea mannoosipitoisuus on pohjimmiltaan vain kaksi N-asetyyliglukosamiinia, joissa on paljon mannoosijäännöksiä, usein melkein yhtä paljon kuin esiaste-oligosakkarideissa on ennen niiden liittämistä proteiiniin.
- Kompleksiset oligosakkaridit ovat saaneet nimensä siksi, että ne voivat sisältää lähes minkä tahansa määrän muun tyyppisiä sakkarideja, mukaan lukien enemmän kuin alkuperäiset kaksi N-asetyyliglukosamiinia.
- Hybridioligosakkaridit sisältävät mannoosijäämiä haaran toisella puolella, kun taas toisella puolella N-asetyyliglukosamiini aloittaa kompleksisen haaran.
Sokerien lisäysjärjestys kasvaviin glykaaniketjuihin määräytyy entsyymien substraattispesifisyyksien ja niiden pääsyn substraattiin niiden liikkuessa eritysreitillä. Näin ollen tämän koneiston järjestäytyminen solun sisällä on tärkeässä asemassa määriteltäessä, mitä glykaaneja valmistetaan.
Golgin entsyymitEdit
Golgin entsyymit ovat avainasemassa määriteltäessä erityyppisten glykaanien synteesiä. Entsyymien toimintajärjestys heijastuu niiden sijaintiin Golgin pinossa:
| entsyymit | sijainti Golgissa |
|---|---|
| Mannosidaasi I | cis-Golgi |
| GlcNAc-transferaasit | mediaalinen Golgi |
| Galaktosyylitransferaasi ja sialyylitransferaasi | trans-Golgi |
Arkeoissa ja prokaryooteissaEdit
Samankaltaisia N-glykaanin biosynteesireittejä on löydetty prokaryooteista ja arkeoista. Verrattuna eukaryootteihin lopullinen glykaanirakenne eubakteereissa ja arkeoissa ei kuitenkaan näytä eroavan paljon endoplasmisessa retikulumissa valmistetusta alkuperäisestä esiasteesta. Eukaryooteissa alkuperäinen esiaste-oligosakkaridi modifioidaan laajasti matkalla solun pinnalle.
Vastaa