” Kuinka suuri on E. coli -solu ja mikä on sen massa?
On 3 joulukuun, 2021 by adminMinkä kokoinen on E. coli -solu ja mikä on sen massa?
Lukutila
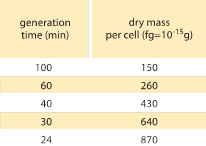
Taulukko 1: Bakteerin massan ja jakautumisajan välinen suhde. Kuivamassa solua kohti on annettu sukupolven (kaksinkertaistumis)ajan funktiona. Massan ehdotetaan kasvavan suunnilleen eksponentiaalisesti kasvunopeuden mukana, kuten alun perin M. Schaechter et al. havaitsivat J. Gen. Microbiol., 19:592, 1958. Solujen kuivapaino laskettiin käyttämällä arvoa 173 µg yhden millilitran OD460-yksikköä kohti (BNID 106437). Käytetty kanta on B/r, jota käytetään yleisesti varhaisissa bakteerien fysiologisissa tutkimuksissa. Arvot on otettu lähteestä F. C. Neidhardt, ”Escherichia coli and Salmonella: Cellular and Molecular Biology”, Vol. 1., Chapter 3, ASM Press, 1996.
Tyypillisen bakteerin, kuten E. colin, koko toimii kätevänä standardiviivoittajana pituusasteikkojen luonnehtimisessa molekyyli- ja solubiologiassa. Valo- ja elektronimikroskooppimittausten sukupolviin perustuva ”nyrkkisääntö” E. coli -solun mitoille on määrittää sen halkaisijaksi noin ≈1µm, pituudeksi ≈2µm ja tilavuudeksi ≈1µm3 (1 fL) (BNID 101788). Muoto voidaan approksimoida pallosylinteriksi eli sylinteriksi, jossa on puolipallonmuotoisia korkkeja. Ilmoitetun halkaisijan ja pituuden perusteella voimme laskea tarkemman arvion tilavuudesta ≈1,3 µm3 (5π/12 tarkalleen ottaen). Tämän arvon ja edellä esitetyn nyrkkisääntöarvon välinen ero osoittaa, kuinka epäjohdonmukaista on, että nyrkkisääntöjä käytettäessä on helppo elää niiden kanssa. Yksi yksinkertaisimmista tavoista arvioida bakteerin massaa on hyödyntää E. coli -solun ≈1 µm3 -tilavuutta ja olettaa, että sen tiheys on sama kuin veden. Tämä naiivi arvio johtaa toiseen vakioarvoon, nimittäin siihen, että E. colin kaltaisen bakteerin massa on ≈1 pg (pico=10-12). Koska useimmat solut koostuvat noin 2/3:sta vettä (BNID 100044, 105482) ja muiden osien, kuten proteiinien, ominaistiheys on noin 1,3 kertaa veden tiheys (BNID 101502, 104272), muunnos solun tilavuudesta massaksi on noin 10 %:n tarkkuudella tarkka.
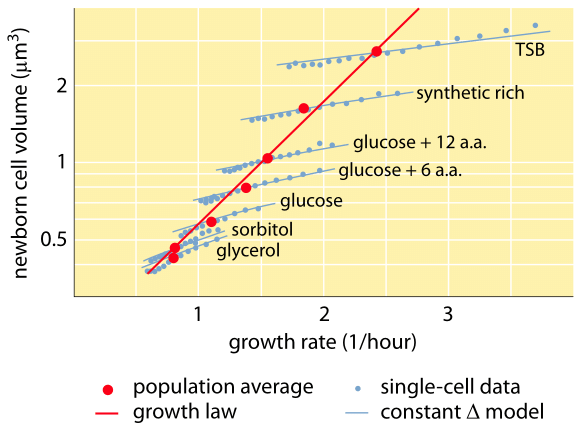
Kuvio 1. Solun tilavuuden muuntaminen massaksi: Solutilavuuden ja kasvunopeuden välinen suhde. Mikroskopian ja mikrofluidisten laitteiden avulla solutilavuutta voidaan mitata yhden solun tasolla eri olosuhteissa, mikä vahvistaa, että keskimääräinen solutilavuus kasvaa eksponentiaalisesti kasvunopeuden myötä. Sitä vastoin solujen välinen vaihtelu tietyissä olosuhteissa skaalautuu eri tavalla. Yksittäisten solujen käyttäytymisen vaihtelua käytetään solujen koon säätelyn mallien testaamiseen. (Mukailtu lähteestä S. Taheri-Araghi et al., Curr. Biol. 25:385, 2015.)
Yksi bakteerien fysiologian klassisista tuloksista korostaa, että solujen ominaisuuksien plastisuus johtuu solumassan riippuvuudesta kasvunopeudesta. Yksinkertaisesti sanottuna nopeampi kasvunopeus liittyy suurempiin soluihin. Tämä havainto viittaa fysiologisiin muutoksiin, joissa kasvunopeutta lisäävät väliaineet tuottavat myös suurempia soluja, kuten kuvassa 1 on esitetty. Tämän havaittiin pitävän paikkansa myös geneettisesti, kun nopeampaan kasvunopeuteen johtaneissa pitkäaikaisissa kokeellisissa evoluutiotutkimuksissa havaittiin suurempia solumääriä (BNID 110462). Tällaiset havainnot auttavat meitä hälventämään myytin ”solusta” – jossa ihmiset, usein tahattomasti, käyttävät yhtä solua koskevia mittaustuloksia tehdäkseen johtopäätöksiä muista solutyypeistä tai samasta solutyypistä eri olosuhteissa. Dennisin ja Bremerin klassisissa tutkimuksissa systematisoitiin nämä mittaukset ja todettiin, että kuivamassa vaihtelee taulukossa 1 esitetyllä tavalla 100 minuutin välein jakautuvien solujen keskiarvosta 148 fg:n arvoon ja 24 minuutin jakautumisajalla jakautuvien solujen arvoon 865 fg:n arvoon, mikä merkitsee yli viisinkertaista eroa kasvunopeudesta riippuen. Samanlainen suuntaus on havaittu muissa organismeissa (esim. orastuvalla hiivalla, BNID 105103). Noin 70-prosenttisessa vedessä nämä arvot vastaavat tilavuudeltaan noin 0,4-2,5 µm3 -aluetta. Miten voimme järkeistää nopeammin kasvavien solujen suuremmat koot? Tästä kysymyksestä keskustellaan vielä tänäkin päivänä (Molenaar D. et al. MSB 5:323, 2009; Amir, A., Phys. Rev, Let., 112:208102, 2014). Selitykset vaihtelevat väitteistä, joiden mukaan se on etulyöntiasemassa resurssien jakamisessa, väitteisiin, joiden mukaan se on itse asiassa vain sivuvaikutus siitä, että solulla on noin 60 minuutin sisäänrakennettu ajanjakso siitä, kun solu päättää, että se on kerännyt tarpeeksi massaa aloittaakseen jakautumisen valmistelut, ja siihen asti, kunnes se saa DNA:n replikaation ja jakautumisen päätökseen. Tämä suunnilleen vakio ”viiveaika” johtaa tässä päättelyssä solun keskimääräisen massan eksponentiaaliseen riippuvuuteen kasvunopeudesta (Amir, A., Phys. Rev, Let.., 112:208102, 2014).
Menetelmät solujen tilavuuden mittaamiseksi vaihtelevat Coulter-laskurin käytöstä ((BNID 100004), joka päättelee tilavuuden perustuen pienen aukon vastuksen muutoksiin solun kulkiessa siinä, ja suorempiin mittauksiin, joissa käytetään fluoresenssimikroskopiaa, joka mittaa solujen pituuksia ja halkaisijoita eri olosuhteissa (kuva 1 ja BNID 106577, 111480). Yllättäen se, että eri laboratoriot eivät aina pääse samoihin arvoihin, voi johtua kalibrointimenetelmien tai tarkkojen kantojen ja kasvuolosuhteiden eroista. Ennennäkemätön kyky mitata solujen massaa saavutetaan punnitsemalla soluja tehokkaasti mikroskooppisen kannattimen päällä. Kuten kuvassa 2A havainnollistetaan, nestevirtauksen avulla solu pakotetaan edestakaisin onttoon kantiiliin. Mittauksessa hyödynnetään sitä, että solun massa vaikuttaa konsolin värähtelytaajuuteen. Tämä taajuus voidaan mitata ilmiömäisellä tarkkuudella, ja sen avulla voidaan päätellä massat femtogrammin tarkkuudella. Kun nesteen virtaussuuntaa muutetaan, solu jää loukkuun minuuteiksi tai pidemmäksi aikaa, ja sen massan kertymisnopeutta mitataan jatkuvasti yksittäisen solun tasolla. Tämän tekniikan ensimmäisessä sovelluksessa osoitettiin, että yksittäiset solut, jotka ovat suurempia, kerryttävät massaa myös nopeammin, mikä valaisee pitkäaikaista kysymystä: onko solujen kasvu lineaarista ajan kanssa vai kuvaako sitä paremmin suunnilleen eksponentiaalinen suuntaus? Erot voivat olla pieniä, mutta näiden vallankumouksellisten mahdollisuuksien avulla nähtiin selvästi, että jälkimmäinen skenaario edustaa paremmin tilannetta useissa testatuissa solutyypeissä, kuten kuvassa 2B on esitetty.
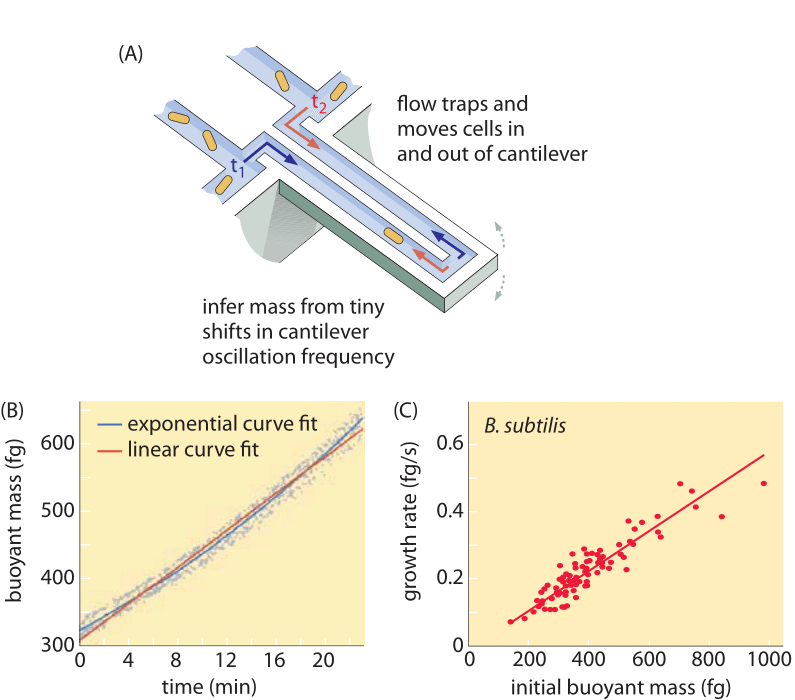
Kuva 2: Kelluvan massan käyttö yksittäisten solujen kasvun mittaamiseen. (A) Mikrometrin kokoinen cantilever värähtelee korkealla taajuudella, ja solujen massa voidaan määrittää värähtelytaajuuden muutoksista. (B) Ajan kuluessa mitattuna tästä saadaan yksittäisen solun massan kertymiskäyrä kuvan mukaisesti. (C) Kuvassa on B. subtilis -soluja. Lineaarisen ja eksponentiaalisen kasvumallin ennusteiden vertailu on esitetty parhaana sovituksena. Samankaltaisuus osoittaa, kuinka lähellä nämä kaksi mallia ovat vain kaksinkertaisen kasvun alueella solusyklin aikana. Solun kuivapaino on noin 4 kertaa kelluva massa. (Mukailtu lähteestä M. Godin et al., Nature Meth. 7:387, 2010.)





Vastaa