Hebbin teoria
On 14 joulukuun, 2021 by admin Arviointi | Biopsykologia | Vertaileva | Kognitiivinen | Kehitys | Kieli | Yksilölliset erot | Persoonallisuus | Filosofia | Yhteiskuntatieteellinen |
Metodit | Tilastot | Tilastot | Kliininen | Kasvatus | Koulutus | Teollisuus | Ammatilliset kohteet | Maailmanlaajuinen psykologian alkeet |
Kognitiivinen psykologia:Hebbin teoria kuvaa synaptisen plastisuuden perusmekanismia, jossa synaptisen tehokkuuden lisääntyminen johtuu siitä, että presynaptinen solu stimuloi postsynaptista solua toistuvasti ja pysyvästi. Donald Hebb esitteli sen vuonna 1949, ja sitä kutsutaan myös nimellä Hebbin sääntö, Hebbin postulaatti ja solujen kokoonpanoteoria, ja siinä todetaan:
Oletetaan, että palautuvan toiminnan (tai ”jäljen”) pysyvyys tai toistuvuus pyrkii aiheuttamaan pysyviä solumuutoksia, jotka lisäävät sen vakautta…. Kun solun A aksoni on tarpeeksi lähellä herättääkseen solun B ja osallistuu toistuvasti tai pysyvästi sen sytyttämiseen, jommassakummassa tai molemmissa soluissa tapahtuu jokin kasvuprosessi tai aineenvaihdunnallinen muutos siten, että A:n tehokkuus yhtenä B:tä sytyttävistä soluista kasvaa.
Teoria tiivistetään usein muotoon ”solut, jotka ampuvat yhdessä, johdottuvat yhteen”, vaikka tämä on hermoston liiallinen yksinkertaistaminen, jota ei pidä ottaa kirjaimellisesti, eikä se myöskään edusta tarkasti Hebbin alkuperäistä lausumaa solujen yhteyksien voimakkuuden muutoksista. Teoriaa käytetään yleisesti selittämään joitakin assosiatiivisen oppimisen muotoja, joissa solujen samanaikainen aktivointi johtaa synaptisen voimakkuuden voimakkaaseen kasvuun. Tällaista oppimista kutsutaan Hebbiläiseksi oppimiseksi.
Hebbiläiset engrammit ja solujen yhdistämisteoria
Hebbiläinen teoria koskee sitä, miten neuronit voivat yhdistyä toisiinsa engrammeiksi. Hebbin teoriat solukokoonpanojen muodosta ja toiminnasta voidaan ymmärtää seuraavasta:
”Yleinen ajatus on vanha, että millä tahansa kahdella solulla tai solujärjestelmällä, jotka ovat toistuvasti aktiivisia samaan aikaan, on taipumus ’assosioitua’ niin, että aktiivisuus yhdessä helpottaa aktiivisuutta toisessa.” (Hebb 1949, s. 70) ”Kun yksi solu toistuvasti avustaa toisen solun laukeamista, ensimmäisen solun aksoni kehittää synaptisia nuppuja (tai suurentaa niitä, jos ne ovat jo olemassa), jotka ovat kosketuksissa toisen solun somen kanssa”. (Hebb 1949, s. 63)
Gordon Allport esittää lisäajatuksia koskien solujen kokoonpanoteoriaa ja sen roolia engrammien muodostumisessa autoassosiaatio-käsitteen mukaisesti, jota kuvataan seuraavasti:
”Jos systeemin syötteet aiheuttavat saman aktiivisuuskuvion toistuvan esiintymisen, tuon kuvion muodostavat aktiivisten elementtien joukot assosioituvat yhä voimakkaammin toisiinsa. Toisin sanoen jokaisella elementillä on taipumus kytkeä kaikki muut elementit päälle ja (negatiivisilla painoilla) kytkeä pois päältä elementit, jotka eivät kuulu kuvioon. Toisin sanoen koko kuvio ”assosioituu itsestään”. Voimme kutsua opittua (auto-assosioitunutta) mallia engrammiksi.” (Hebb 1949, s. 44)
Hebbin teoria on ollut ensisijainen perusta perinteiselle näkemykselle, jonka mukaan kokonaisvaltaiselta tasolta analysoituna engrammit ovat neuroniverkkoja tai hermoverkkoja.
Eric Kandelin laboratoriossa tehdyssä työssä on saatu todisteita Hebbin oppimismekanismien osallisuudesta synapseissa merikotilolla Aplysia californica.
Kokeet selkärankaisten keskushermoston synapsien Hebbiläisistä synapsien muutosmekanismeista ovat paljon vaikeammin kontrolloitavissa kuin kokeet suhteellisen yksinkertaisilla ääreishermoston synapseilla, joita on tutkittu meren selkärangattomilla. Suuri osa selkärankaisten neuronien välisiä pitkäaikaisia synaptisia muutoksia (kuten pitkäaikaispotentiaatio) koskevasta työstä edellyttää aivosolujen ei-fysiologista kokeellista stimulointia. Jotkin selkärankaisten aivoissa tutkituista fysiologisesti merkityksellisistä synapsien muutosmekanismeista näyttävät kuitenkin olevan esimerkkejä Hebbin prosesseista. Eräässä tällaisessa tutkimuksessa tarkastellaan tuloksia kokeista, jotka osoittavat, että fysiologisesti relevantti synaptinen aktiivisuus, joka toimii sekä Hebbiläisten että muiden kuin Hebbiläisten mekanismien kautta, voi saada aikaan pitkäkestoisia muutoksia synaptisten voimakkuuksien voimakkuudessa
Periaatteet
Keinotekoisten neuronien ja keinotekoisten hermoverkkojen näkökulmasta Hebbin periaate voidaan kuvata menetelmänä, jonka avulla voidaan määritellä, miten mallineuronien välisiä painoja voidaan muuttaa. Kahden neuronin välinen paino kasvaa, jos neuronit aktivoituvat samanaikaisesti – ja pienenee, jos neuronit aktivoituvat erikseen. Solmuilla, joilla on taipumus olla samanaikaisesti joko molemmat positiivisia tai molemmat negatiivisia, on voimakkaat positiiviset painot, kun taas solmuilla, joilla on taipumus olla vastakkaisia, on voimakkaat negatiiviset painot.
Tämä alkuperäinen periaate on ehkä yksinkertaisin painojen valinnan muoto. Vaikka tämä tarkoittaa, että se voidaan suhteellisen helposti koodata tietokoneohjelmaan ja käyttää verkon painojen päivittämiseen, se myös rajoittaa Hebbian-oppimisen sovellusten määrää. Nykyään termi Hebbin oppiminen viittaa yleensä jonkinlaiseen matemaattiseen abstraktioon Hebbin esittämästä alkuperäisestä periaatteesta. Tässä mielessä Hebbiläisessä oppimisessa oppimissolmujen välisiä painoja säädetään siten, että kukin paino edustaa paremmin solmujen välistä suhdetta. Sellaisenaan monia oppimismenetelmiä voidaan pitää luonteeltaan jossain määrin Hebbiläisinä.
Seuraavassa on kaavamainen kuvaus Hebbiläisestä oppimisesta: (huomaa, että monet muutkin kuvaukset ovat mahdollisia)
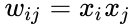
missä 
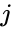
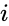
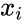
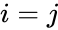
Toinen kaavamainen kuvaus on:
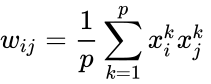
missä 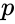

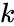
Hebbiläisen oppimisen muunnelma, joka ottaa huomioon estymisen kaltaiset ilmiöt ja monet muut neuraaliset oppimisilmiöt, on Harry Klopfin matemaattinen malli. Klopfin malli toistaa hyvin monia biologisia ilmiöitä, ja se on myös yksinkertainen toteuttaa.
Yleistäminen ja stabiilisuus
Hebbin sääntö yleistetään usein muotoon
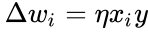
tai 
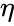
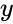
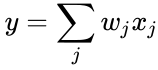
ja edellisen kappaleen yksinkertaistuksessa sekä oppimisnopeus että tulopainot otetaan arvoksi 1. Tämä versio säännöstä on selvästi epävakaa, sillä missä tahansa verkossa, jossa on dominoiva signaali, synaptiset painot kasvavat tai pienenevät eksponentiaalisesti. Voidaan kuitenkin osoittaa, että missä tahansa neuronimallissa Hebbin sääntö on epävakaa. Siksi neuronien verkkomalleissa käytetään yleensä muita oppimisteorioita, kuten BCM-teoriaa, Ojan sääntöä tai yleistettyä Hebbin algoritmia.
Katso myös
- Anti-Hebbiläinen oppiminen
- BCM-teoria
- Sattuman havaitseminen neurobiologiassa
- Dalen periaate
- Yleistetty Hebbiläinen algoritmi
- Ojan periaate
- Ojan periaate
- Muisti
- Metaplastisuus
- Neuraaliset verkostot
- Oja oppimissääntö
- Tetaninen stimulaatio
- Spike-timing-riippuvainen plastisiteetti
- Synaptotrooppinen hypoteesi
Pitkä.term potentiation
- Shouval, Harel Aivojen fysiikka. Oppimisen ja muistin synaptinen perusta: Teoreettinen lähestymistapa. University of Texas Health Science Center at Houston. URL-osoite haettu 2007-11-14.
Lisätietoja
- Hebb, D.O. (1949), The organization of behavior, New York: Wiley
- Hebb, D.O. (1961). ”Distinctive features of learning in the higher animal” J. F. Delafresnaye (Ed.) Brain Mechanisms and Learning, London: Oxford University Press.
- Hebb, D.O., and Penfield, W. (1940). Ihmisen käyttäytyminen otsalohkojen laajamittaisen molemminpuolisen poiston jälkeen. Archives of Neurology and Psychiatry 44: 421-436.
- Allport, D.A. (1985). ”Hajautettu muisti, modulaariset järjestelmät ja dysfasia” Newman, S.K. ja Epstein, R. (toim.) Current Perspectives in Dysphasia, Edinburgh: Churchill Livingstone.
- Bishop, C.M. (1995). Neural Networks for Pattern Recognition, Oxford: Oxford University Press.
- Paulsen, O., Sejnowski, T. J. (2000). Luonnolliset aktiivisuusmallit ja pitkäaikainen synaptinen plastisuus. Current opinion in neurobiology 10 (2): 172-179.
- Yleiskatsaus
- Hebbiläisen oppimisen opetusohjelma (Osa 1: Uutuuksien suodatus, Osa 2: PCA)
|
v-d-e
Neuroetiikka |
||
|---|---|---|
| Käsitteitä neuroetiikassa |
Feedforward – Sattumanilmaisin – – Ympäristö – Vaisto – Ominaisuusilmaisin – Keskuskuvion generaattori (CPG) -NMDA-reseptori – Lateraalinen inhibitio – Kiinteä toimintamalli – Kroghin periaate-Hebbin teoria- Äänen lokalisointi |
200px
|
| NEUROETOLOGIAN HISTORIA |
Theodore Holmes Bullock – Walter Heiligenberg -Niko Tinbergen- Konrad Lorenz- Eric Knudsen-Donald Griffin – Donald Kennedy – Karl von Frisch – Erich von Holst – Jörg-Peter Ewert |
|
| Methods in Neuroethology |
Whole Cell Patch Clamp |
|
| Model Systems in Neuroethology |
Animaalisten kaikuluotaus – Vaggle- Dance- Electric Fish – Vision in toads – Neuroethology of Frog Audition – Infrared sensing in snakes |
|
Tällä sivulla käytetään Wikipedian Creative Commons -lisensoitua sisältöä (Näytä tekijät).
Vastaa