Frontiers in Psychology
On 28 syyskuun, 2021 by adminCorpus callosum (CC) on ensisijainen aivopuoliskoja yhdistävä yhdysside. Näin ollen, vaikka termi kommissurotomia ei ole virheellinen kuvaamaan niin sanottua ”halkaistua aivoa”, ja se on täsmälleen oikea alkuperäisistä ihmisen kirurgisista operaatioista, joissa kaikki kommissuuret katkaistiin (Gazzaniga et al., 1967), myöhemmissä leikkauksissa katkaistiin vain CC, jolloin käyttöön tuli termi ”callosotomia”, joka on edelleen asianmukaisempi termi näissä tapauksissa (esim. Franz ym., 1996, 2000).
Evoluution kannalta CC on viimeaikainen kehityskulku, joka on havaittu vain istukkanisäkkäillä (Katz ym., 1983; Mihrshahi, 2006). CC:n tarjoaman toiminnallisuuden uskotaan edeltävän ainakin osittain anteriorista kommissuuraa (anterior commissure, AC), joka on havaittu ei-placentaalisilla nisäkkäillä, kuten pussieläimillä (Heath ja Jones, 1971; Ashwell ym., 1996). Vaikka ihmisen aivoissa on edelleen joitakin AC:n jäännösreittejä, sen kytkeytyvyyttä ja rakennetta ei ole tutkittu riittävästi nykyisessä kirjallisuudessa, ja harvat havainnot ovat melko epäjohdonmukaisia (Lasco et al., 2002; Patel et al., 2010; Choi et al., 2011). Esimerkiksi magneettikuvaukseen (MRI) perustuvat havainnot ovat ristiriitaisia sen suhteen, onko AC samankokoinen naisilla ja miehillä (Lasco et al., 2002; Patel et al., 2010). Suhteellisen tuoreessa tutkimuksessa, jossa käytettiin in vivo -tekniikkaa, diffuusiotensorikuvantamista (DTI), jossa hyödynnetään vesimolekyylien diffuusiota aksoneissa, Patel ja muut (2010) tutkivat kahdeksaa koehenkilöä ja eristivät onnistuneesti AC:n kuidut viideltä koehenkilöltä. Löydösten joukossa oli yhdellä koehenkilöllä suuri osa AC:n takaosan kuiduista, jotka kulkivat molemmin puolin parietaalisille alueille, missä suurin osa niistä päättyi, ja pienempi nippu päättyi precentraaliseen gyrusiin, ja vielä yksi nippu meni ohimolohkoon. Tällaisia yksilöllisiä eroja havaittiin kaikilla osallistujilla, mutta suurin osa koehenkilöistä osoitti kahdenvälisiä projektioita orbitofrontaaliseen, parietaaliseen ja temporaaliseen aivokuoreen, joihin CC ei selvästi projisoi. Yhdessä havainnot viittaavat siihen, että AC: llä on sekä primaarisia että vestigiaalisia projektioita neokortikaalisille alueille, joihin CC ei projisoi, kuten muutkin ovat ehdottaneet (Di Virgilio et ai., 1999; Patel et ai., 2010).
Näkemyksemme mukaan AAA tukeutuu voimakkaasti neokortikaalisiin verkostoihin ja yhteyteen subkortikaalisten rakenteiden, kuten basaaliganglioiden ja talamuksen, kuten basaaliganglioiden ja talamuksen, kanssa (muun muassa muiden muassa, joita ei ole käsitelty ehdotuksessamme erityisesti) (Franz, 2012). Näin ollen kaikilla AC:hen liittyvillä evoluutiokehityksillä saattaa olla jonkinlainen rooli myös AAA-mallissa. Löysimme kuitenkin vain vähän kirjallisuutta, jossa käsitellään erityisesti tätä mahdollista yhteyttä.
AC:n roolia on tutkittu tehtävissä, jotka todennäköisesti vaativat huomion muotoja, kuten visuaalisen huomion tehtäviä. On esimerkiksi tunnettua, että acallosalipopulaatioita on tutkittu visuaalista integraatiota sisältävien tehtävien yhteydessä (Gazzaniga et al., 1962, 1965; Sperry, 1968; Levy et al., 1972). Viime aikoina Corballis (1995) pyrki käsittelemään visuaalista integraatiota ja tarkkaavaisuutta selittääkseen aiempia ristiriitaisuuksia acallosal-tutkimuksessa. Corballis ehdotti kaksijakoista visuaalista tarkkaavaisuusjärjestelmää; yksi osa (joka koskee muotoa) on automaattinen ja lokalisoitu kummallekin aivopuoliskolle, ja toinen osa (joka koskee liikettä ja sijaintia) on vapaaehtoinen ja subkortikaalinen. AAA-malli voi perustua tähän havaintoon ja puuttua kirjallisuudessa havaittuihin ristiriitoihin. Ehdotetun mallin perusteella useilla huomion tasoilla on muitakin reittejä kuin CC, jotka saattavat välittää tiedonsiirtoa aivopuoliskojen välillä. Yksi mahdollisuus liittyy basaaliganglion piiriin, jota ehdotamme, että se on mahdollisesti kriittinen huomion jakamisen välittämisessä aivopuoliskojen välillä (Franz, 2012). Kun CC leikataan, suora huomion siirto aivokuoren väylien kautta poistuu suurelta osin (ellei AC:llä ole tietenkään vararoolia). Visuaalisen informaation tarkkaavaisuuteen perustuvaa integrointia voitaisiin kuitenkin edelleen välittää subkortikaalisten ratojen ja/tai AC:n kautta. Tällä tavoin Corballisin (1995) havaitsema kaksiosainen tarkkaavaisuusjärjestelmä voisi olla läsnä, koska tarkkaavaisuuteen perustuva visuaalisen integraation välittäminen tapahtuu AC:n kautta (ja/tai basaaliganglion ja siihen liittyvien piirien kautta: Franz, 2012).
Näköä ja tarkkaavaisuutta koskevia tutkimuksia ei tarvitse tarkastella täysin erillään motorista toimintaa säätelevästä järjestelmästä. Omien havaintojemme (Franz, 2004; Franz ja Packman, 2004) perusteella huomion eri muodot (visuaalinen ja sisäinen-nonvisuaalinen) liittyvät läheisesti motoriseen toimintaan, ja on perusteltua, että mahdollinen avain AC:n mahdollisen huomioroolin ymmärtämiseen on toiminnan, näön ja huomion välisessä yhteydessä. Aikaisemmassa tutkimuksessa Franz (2004) sai normaalit osallistujat piirtämään ympyröitä bimanuaalisesti erilaisilla visuaalisen palautteen tai sisäisen huomion manipuloinneilla kumpaankin käteen. Sisäisen huomion muotoa saavan käden piirtämien ympyröiden koko muuttui suuremmaksi tavalla, joka oli samankaltainen kuin vaikutukset, joita esiintyi kyseisen käden visuaalisen palautteen läsnä ollessa (suhteessa siihen, ettei huomiota / visuaalista palautetta ollut). AAA-mallimme mukaan huomio on keskeinen komponentti motorisen tuotoksen säätelyssä (sekä sen valinnassa, mitä motorista toimintaa suunnitellaan), ja sikäli kuin nykyiset todisteet osoittavat, emme voi sulkea pois sitä, että AC:llä on rooli tässä toiminnassa. AC:n roolin suora testaaminen missä tahansa kognitiivisessa tehtävässä ei kuitenkaan ole triviaalia.
Testaus siitä, osallistuuko AC kognitiivisiin toimintoihin, on neuropsykologisessa tutkimuksessa lähes mahdotonta, koska osallistujia, joilla on paikallinen AC, ei ole helppo löytää (emmekä tiedä yhtään). Siksi saattaisi olla paljon intuitiivisempaa arvioida neurologisia eroja normaalissa AC:ssä ja korreloida neurologiset tiedot käyttäytymistietojen kanssa. Esitettyä traktografiamenetelmää (AC:n selvittäminen) voidaan soveltaa suhteellisen helposti suuriin näytteisiin normaaliväestöstä, ja ihanteellisessa tapauksessa se voidaan yhdistää näiden samojen henkilöiden suorituskykyyn (joka perustuu tehtäviin, jotka on suoritettu DTI-kokoelmasta erillisten testausistuntojen aikana) käyttämällä tarkoituksenmukaisesti suunniteltuja kognitiivisia tehtäviä. Tässä osoitamme, että DTI voidaan suorittaa ja että siitä voidaan saada käyttökelpoisia tuloksia pienellä otoksella neurologisesti normaaliväestöön kuuluvia henkilöitä. Yleinen ehdotus olisi suorittaa samanlaisia menetelmiä suuremmalla otoskoolla ja (kuten edellä ehdotettiin), korreloida havainnot (esimerkiksi AC:n tiheyden proxy-mittauksesta) kognitiivisen tehtävän muuttujien kanssa.
Kuvassa 1 näytämme DTI-seurannan tulokset 10 naispuoliselta osallistujalta saaduista tiedoista, jotka on saatu Johns Hopkinsin yliopiston julkisesta DTI-tietokannasta (ks. Kiitokset täydellisen lähteen löytämiseksi). Keski-ikä oli 22 vuotta (SD = 2,5). Kuvat käsiteltiin FSL Diffusion Toolboxilla (Smith et al., 2004; Woolrich et al., 2009; Jenkinson et al., 2012), ja menettelyissä käytettiin niitä menettelyjä, joita on käytetty aiemmissa tutkimuksissa muilla kuitupoluilla (FDT; Behrens et al., 2003a,b, 2007; Johansen-Berg et al., 2004). Lyhyesti sanottuna kuvat yhdistettiin ensin 4d-tilavuudeksi kunkin osallistujan osalta ja kallot kuorittiin FSL:n Brain Extraction Tool -työkalulla (BET; Smith, 2002; Jenkinson et al., 2005). Poistetut kuvat korjattiin pään vähäisen liikkeen ja kentän suunnan muutosten aiheuttamien pyörrevirtojen osalta. Diffuusiotensorit sovitettiin tuottamaan kuvia, joissa näkyvät ensisijaiset diffuusiosuuntaukset ja FA-tasot kussakin vokselissa, jotta niitä voitiin käyttää värikarttoina maskeerauksen aikana. Diffuusioparametrien bayesiläinen estimointi, joka saatiin käyttämällä näytteenottotekniikoita ristikkäisten kuitujen kanssa (BEDPOSTX; ”X” viittaa ristikkäisiin kuituihin), suoritettiin diffuusion koon ja suunnan jakauman luomiseksi ottaen huomioon kaksi ristikkäistä kuitua, jotta myöhemmin voitiin suorittaa traktografia diffuusiovokselien yli.
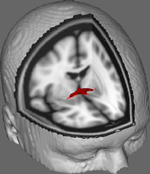
KUVA 1. Diffuusioparametrit. Keskiarvoistetut jäljitystulokset 10:llä kontrolliosallistujalla päällekkäin vakiomuotoisen T1-painotetun MRI-kuvan kanssa, jossa on 2 mm:n isotrooppiset vokselit anatomista referenssiä varten.
Keskiarvoistetut jäljitystulokset kaikilla 10:llä kontrolliosallistujalla päällekkäin vakiomuotoisen T1-painotetun MRI-kuvan kanssa, jossa on 2 mm:n isotrooppiset vokselit anatomista referenssiä varten. Ryhmän keskimääräinen FA-suhde oli 0,3146 (SD = 0,0472). Nämä traktografiatulokset paljastavat, mikä on tärkeää, että AC: n rakennetta voidaan selvittää, ja siksi sillä on potentiaalia jatkotutkimuksiin, joissa korreloidaan käyttäytymistehtävien suorituskykyarvoja FA-arvojen kanssa käyttämällä suurempaa otoskokoa.
Yhteenvetona voidaan todeta, että tällä hetkellä on olemassa niukasti kirjallisuutta, eikä käytettävissä olevasta kirjallisuudesta yksikään ei nimenomaisesti liitä AC: tä prosesseihin, jotka liittyvät aiempaan kohdeartikkelimme kohteena olevaan AAA-malliin (Franz, 2012). Käyttämällä tässä ehdotetun kaltaista menettelyä on mahdollista testata suurempia otoskokoja normaaliväestöstä ja korreloida DTI-tulosten FA-arvot (esimerkiksi) tietyn riippuvaisen muuttujan kanssa, joka saadaan tarkoituksenmukaisesti kehitetystä tarkkaavaisuustehtävästä (jonka menettelyt ovat käynnissä oleva kehitys laboratoriossamme).
Irintaristiriitojen selvitys
Tekijät ilmoittavat, että tutkimus suoritettiin ilman kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Kiitokset
Kiitämme kiitollisena Human Brain Projectia ja kansallista tutkimusresurssikeskusta (USA) ja John Hopkinsin yliopistoa siitä, että he sallivat DTI-tietojensa julkisen käytön. Olemme myös kiitollisia Uuden-Seelannin kuninkaallisen seuran Marsden Fundin anteliaasta apurahatuesta (Elizabeth A. Franzille).
Corballis, M. (1995). Visuaalinen integraatio jakautuneissa aivoissa. Neuropsychologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). Huomion kohdistaminen tavoiteohjattujen toimien oppimiseen: kognitiivisen neurotieteen kehys, jossa keskitytään basaaliganglioihin. Front. Psychol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., and Jones, E. (1971). Interhemispheric pathways in the absence of a corpus callosum. Kokeellinen tutkimus commissural connexions in the marsupial phalanger. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
Vastaa