Bakteerisolujen rakenne ja toiminta
On 10 lokakuun, 2021 by adminBakteerisolujen rakenne ja toiminta (sivu 5)
(Tässä luvussa on 10 sivua)
© Kenneth Todar, PhD
Soluseinä
Soluseinät
Bakteerien soluseinät ansaitsevat erityishuomiota useista syistä:
1. Ne ovat elinkelpoisuuden kannalta olennainen rakenne, kuten edellä on kuvattu.
2. Ne koostuvat ainutlaatuisista komponenteista, joita ei löydy mistään muualta luonnosta.
3. Ne ovat yksi tärkeimmistä antibioottien hyökkäyskohteista.
4. Ne tarjoavat ligandeja kiinnittymistä varten ja reseptoripaikkoja lääkeaineille tai viruksille.
5. Ne ovat tärkeä osa elinkykyä. Ne aiheuttavat eläimille tautioireita.
6. Ne mahdollistavat bakteerikantojen immunologisen erottelun ja immunologisen vaihtelun.
Useimmilla prokaryooteilla on jäykkä soluseinä.Soluseinä on olennainen rakenne, joka suojaa soluprotoplastia mekaanisilta vaurioilta ja osmoottiselta repeytymiseltä tai lyysiltä.Prokaryootit elävät yleensä suhteellisen laimeissa ympäristöissä siten, että liukoisten aineiden kertyminen prokaryoottisolun sytoplasman sisälle ylittää huomattavasti ulkoisen ympäristön kokonaispitoisuuden. Näin ollen plasmakalvon sisäpuolelle kohdistuva osmoottinen paine voi olla 10-25 atm. Koska kalvo on herkkä ja plastinen rakenne, sitä on rajoitettava huokoisesta ja jäykästä materiaalista valmistetulla ulkoseinällä, jolla on suuri vetolujuus. Tällainen materiaali on mureiini, joka on bakteerien soluseinien kaikkialle levinnyt komponentti.
Mureiini on ainutlaatuinen peptidoglykaanityyppi, joka on lyhyiden aminohappoketjujen (peptidien) ristisilloittamien disakkaridien (glykaanien) polymeeri. Peptidoglykaaneja on monenlaisia. Kaikki bakteeriperäiset peptidoglykaanit sisältävät N-asetyylimuramiinihappoa, joka on mureiinin määritelmäkomponentti. Arkeoiden soluseinät voivat koostua proteiineista, polysakkarideista tai peptidoglykaanin kaltaisista molekyyleistä, mutta ne eivät koskaan sisällä mureiinia. Tämä ominaisuus erottaa bakteerit arkeista.
Gram-positiivisissa bakteereissa (bakteereissa, jotka säilyttävät purppuranpunaista kristalliviolettiväriainetta Gram-värjäysmenetelmässä) soluseinämä koostuu useista peptidoglykaanikerroksista. Peptidoglykaanikerroksia vastaan kohtisuoraan kulkee ryhmä molekyylejä, joita kutsutaan teichoiinihapoiksi ja jotka ovat ainutlaatuisia grampositiivisille soluseinille (kuva 14). 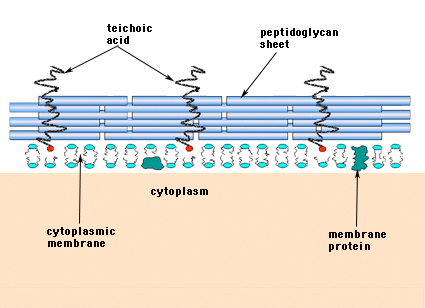
Kuva 14. Grampositiivisten bakteerien soluseinän rakenne. Seinämä onsuhteellisen paksu, ja se koostuu monista peptidoglykaanikerroksista, joiden välissä on teichoehappoja, jotka kulkevat kohtisuoraan peptidoglykaanilevyihin nähden.
Gramnegatiivisissa bakteereissa (jotka eivät säilytä kristalliviolettia) soluseinä koostuu yhdestä peptidoglykaanikerroksesta, jota ympäröi kalvorakenne, jota kutsutaan ulkokalvoksi.Gramnegatiivisten bakteerien ulkokalvo sisältää poikkeuksetta yhden ainoan komponentin, lipopolysakkaridin (LPS tai endotoksiini), joka on myrkyllistä eläimille. Gramnegatiivisilla bakteereilla ulkokalvon ajatellaan yleensä olevan osa soluseinää (kuva15).

Kuva15. Gramnegatiivisen soluseinän rakenne. Seinämä on suhteellisenohut ja sisältää paljon vähemmän peptidoglykaania kuin grampositiivinen seinämä.myös teiköhappoja ei ole. Gramnegatiivinen soluseinä koostuu kuitenkin ulkokalvosta, joka on peptidoglykaanikerroksen ulkopuolella. Ulkokalvo on kiinnittynyt peptidoglykaanikerrokseen ainutlaatuisella lipoproteiinimolekyylien ryhmällä.
Gram-positiivisissa bakteereissa soluseinä on paksu (15-80nanometriä) ja koostuu useista peptidoglykaanikerroksista. Gramnegatiivisissa bakteereissa soluseinämä on suhteellisen ohut (10 nanometriä) ja koostuu yhdestä peptidoglykaanikerroksesta, jota ympäröi ulkokalvo.
E.colin peptidoglykaanirakenne ja -järjestys edustaa kaikkia Enterobacteriaceae-bakteereja sekä monia muita gramnegatiivisia bakteereja. Glykaaninen selkäranka koostuu N-asetyyliglukosamiinin (G) ja N-asetyylimuramiinihapon (M) vuorottelevista molekyyleistä, jotka on yhdistetty beeta-1,4-glykosidisidoksella. N-asetyylimuramiinihapon (M) 3-hiili on korvattu pyruviatista peräisin olevalla laktyylieetteriryhmällä. Laktyylieetteri yhdistää glykaanirungon peptidin sivuketjuun, joka sisältää L-alaniinia (L-ala), D-glutamaattia (D-glu), diaminopimeliinihappoa (DAP) ja D-alaniinia (D-ala). MurNAc on ainutlaatuinen bakteerien soluseinissä, samoin kuin D-glu, DAP ja D-ala. E. colin muramiinihapon alayksikkö on esitetty seuraavassa kuvassa16.

Kuva 16. Escherichia coli -bakteerin peptidoglykaanin muramiinihappo-alayksikön rakenne.Tätä mureiinityyppiä esiintyy useimmissa gramnegatiivisissa bakteereissa. Glykaanirunko on kahden aminosokerin, N-asetyyliglukosamiinin (G) ja N-asetyylimuramiinihapon (M), toistuva polymeeri. N-asetyyli-muramiinihappoon on kiinnittynyt trapeptidi, joka koostuu L-ala-D-glu-DAP-D-ala:sta. b. Muramiinihappo-alayksikön lyhennetty rakenne. c. Muramiinihappo-alayksikkö. Läheiset tetrapeptidisivuketjut voivat olla sidoksissa toisiinsa yhden ketjun DAP:n ja toisen ketjun D-ala:n välisellä interpeptidisidoksella. d. Molekyylin polymeerinen muoto.
Mureiinin säikeet kootaan periplasmassa noin 10:stä muramiinihapon alayksiköstä. Tämän jälkeen säikeet liitetään toisiinsa muodostaen jatkuvan glykaanimolekyylin, joka ympäröi solun.Jos niiden läheisyys sen sallii, glykaanin selkärangasta ulkonevat tetrapeptidiketjut voidaan ristisilloittaa DAP:n vapaan aminoryhmän ja läheisen D-ala:n vapaan karboksiryhmän välisellä interpeptidisidoksella.Peptidoglykaanin kokoaminen plasmakalvon ulkopuolelle tapahtuu periplasmisen entsyymiryhmän, transglykosylaasien, transpeptidaasien ja karboksypeptidaasien, välityksellä. Penisilliinin ja siihen liittyvien beetalaktaamiantibioottien vaikutusmekanismi on transpeptidaasi- ja karboksypeptidaasientsyymien estäminen mureiinisoluseinän kokoamisessa. Näin ollen betalaktamiantibioottien sanotaan ”estävän soluseinän synteesin” bakteereissa.
Peptidoglykaanimolekyylin glykaanista selkärankaa voi pilkkoa lysotsyymi-niminen entsyymi, jota on eläinserumissa, kudoksissa ja eritteissä sekä fagosyyttisessä lysosomissa. Lysotsyymin tehtävänä on pilkkoa bakteerisoluja konstitutiivisena puolustuskeinona bakteeripatogeeneja vastaan. Jotkin grampositiiviset bakteerit ovat hyvin herkkiä lysotsyymille, ja entsyymi on varsin aktiivinen pieninä pitoisuuksina.Kyynelnesteet (kyyneleet) voidaan laimentaa 1:40 000:een ja ne säilyttävät kykynsä lyysiä tiettyjä bakteerisoluja. Gramnegatiiviset bakteerit ovat vähemmän alttiita lysotsyymin hyökkäykselle, koska niiden peptidoglykaanin suojana on ulkokalvo. Lysotsyymin tarkka pilkkomiskohta on N-asetyylimuramiinihapon (M) ja N-asetyyliglukosamiinin (G) välinen beeta 1,4-sidos siten, että kuvassa 16(a) esitetty muramiinihappo-alayksikkö on tulosta lysotsyymin vaikutuksesta bakteerien peptidoglykaaneihin.
Gram-positiivisissa bakteereissa peptidoglykaanien joukossa on lukuisia erilaisia peptidijärjestyksiä. Parhaiten tutkittu on Staphylococcus aureuksen rakenne, joka on esitetty alla olevassa kuvassa 17. DAP:n (E. coli -bakteerissa) tilalla on diaminohappo L-lysiini (L-lys), ja interpeptidisidoksen (gramnegatiivisissa bakteereissa) tilalla on aminohappojen välinen aminohapposilta, joka yhdistää lysiinin vapaan aminoryhmän läheisen tetrapeptidin sivuketjun D-ala:n vapaaseen karboksiryhmään. Tämä järjestely mahdollistaa ilmeisesti tiheämmät ristisidokset läheisten tetrapeptidin sivuketjujen välillä. S. aureuksessa peptidien välinen silta on peptidi, joka koostuu viidestä glysiinimolekyylistä (ns. pentaglysiinisilta). Beetalaktaamiantibiootit estävät interpeptidisillan muodostumista Gram-positiivisessa mureiinissa samalla tavalla kuin interpeptidisidoksen muodostumista Gram-negatiivisessa mureiinissa. Grampositiiviset bakteerit ovat herkempiä penisilliinille kuin gramnegatiiviset bakteerit, koska peptidoglykaania ei suojaa ulkokalvo ja se on runsaampi molekyyli. Grampositiivisissa bakteereissa peptidoglykaanit voivat vaihdella tetrapeptidin asemassa 3 olevan DAP:n tai L-lys:n tilalla olevan aminohapon ja interpeptidisillan tarkan koostumuksen suhteen. Grampositiivisissa bakteereissa esiintyy ainakin kahdeksan erilaista peptidoglykaanityyppiä.
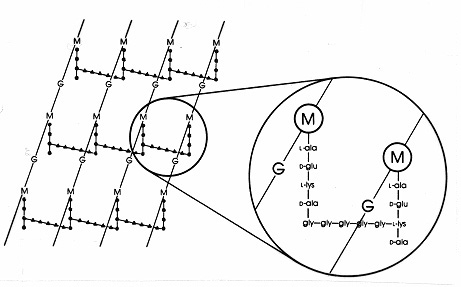
Kuva 17. Staphylococcus aureuksen peptidoglykaanilevyn kaaviokuva. G = N-asetyyli-glukosamiini;M = N-asetyyli-muramiinihappo; L-ala = L-alaniini; D-ala = D-alaniini; D-glu = D-glutamiinihappo; L-lys = L-lysiini. Tämä on yksi grampositiivisissa bakteereissa esiintyvä mureiinityyppi. Verrattuna E. coli -peptidoglykaaniin (kuva7) tetrapeptidissä on L-lys DAP:n (diaminopimeliinihappo) sijasta.L-lys:n vapaa aminoryhmä on korvattu glysiinipentapeptidillä (gly-gly-gly-gly-gly-gly-gly-), josta tulee sitten peptidien välinen silta, joka muodostaa linkin viereisen tetrapeptidin sivuketjussa olevaan D-ala:sta peräisin olevaan karboksiryhmään. Grampositiiviset peptidoglykaanit eroavat toisistaan lajeittain pääasiassa tetrapeptidiketjun kolmannessa asemassa olevien aminohappojen ja interpeptidisillan aminohappokoostumuksen suhteen.
Gramnegatiivisilla bakteereilla voi olla soluseinässään vain yksi yksimolekyylinen mureiinikerros, kun taas grampositiivisilla bakteereilla uskotaan olevan useita kerroksia tai ”kääreitä” peptidoglykaania. Grampositiivisten bakteerien peptidoglykaanikerroksiin liittyy läheisesti ryhmä molekyylejä, joita kutsutaan teiköhapoiksi. Teiköhapot ovat lineaarisia polyglyserolin tai polyribitolin polymeerejä, jotka on korvattu fosfaateilla ja muutamilla aminohapoilla ja sokereilla. Teiköhappopolymeerit ovat toisinaan ankkuroituneet plasmakalvoon (kutsutaan lipoteiköhapoksi, LTA), ja ne ovat ilmeisesti suuntautuneet ulospäin suorassa kulmassa peptidoglykaanikerroksiin nähden. Teichoehapon tehtäviä ei tunneta. Ne ovat välttämättömiä GRAM-positiivisten bakteerien elinkelpoisuuden kannalta luonnossa. Yksi ajatus on, että ne muodostavat kanavan, jossa on säännönmukaisesti suuntautuneita negatiivisia varauksia, joiden kautta positiivisesti varautuneet aineet kulkevat monimutkaisen peptidoglykaaniverkoston läpi. Toisen teorian mukaan teichoiinihapot osallistuvat jollakin tavalla muramiinihappoyksiköiden säätelyyn ja kokoamiseen plasmakalvon ulkopuolella. Erityisesti streptokokkeissa on tapauksia, joissa teiköhappojen on todettu olevan osallisena bakteerien tarttumisessa kudospintoihin.
luku jatkuu
Edellinen sivu
.
Vastaa