Estrutura e Função das Células Bacterianas
On Outubro 10, 2021 by adminEstrutura e Função das Células Bacterianas (página 5)
(Este capítulo tem 10 páginas)
© Kenneth Todar, PhD
Muralha de Células
Os celas de bactérias merecem atenção especial por várias razões:
1. Eles são uma estrutura essencial para a viabilidade, como descrito acima.
2. Eles são compostos de componentes únicos encontrados em nenhum outro lugar na natureza.
3. Eles são um dos locais mais importantes para o ataque de antibióticos.
4. Eles fornecem ligandos para a aderência e receptores de drogas ou vírus.
5. Causam sintomas de doença em animais.
6. Fornecem distinção imunológica e variação imunológica entre estirpes de bactérias.
A maioria dos procariotas tem parede celular rígida. A parede celular é uma estrutura essencial que protege o proto-plastômero celular dos danos mecânicos e da ruptura osmótica ou lise.A parede celular é uma estrutura essencial que protege o citoplasma de células procariotas de danos mecânicos e da ruptura osmótica ou lise. Assim, a pressão osmótica contra o interior da membrana plasmática pode ser o equivalente a 10-25atm. Uma vez que a membrana é uma estrutura plástica delicada, ela deve ser contida por uma parede externa feita de material poroso, rígido e com alta resistência à tração. Tal material é a mureína, o componente ubíquo das paredes celulares bacterianas.
Mureína é um tipo único depeptidoglicano, um polímero de dissacarídeos (glicano) reticulado por ácidos de cadeia curta (peptídeo). Existem muitos tipos de peptidoglicanos. Todos os Bacterialpeptidoglicanos contêm ácido N-acetilmurâmico, que é o componentedefinitivo da mureína. As paredes celulares da Archaea podem tornar-se compostas de proteínas, polissacarídeos, ou moléculas semelhantes a peptidoglicanos, mas nunca contêm mureína. Esta característica distingue as Bactérias da Archaea.
Inthe Gram-positiveBacteria (aquelas que retêm o corante violeta de cristal roxo quando submetidas ao procedimento de coloração de Gram), a parede celular consiste em várias camadas de peptidoglicano. O perpendicular às folhas de peptidoglicano é um grupo de moléculas chamado ácido teicóico que são exclusivas da parede celular Gram-positiva (Figura 14). 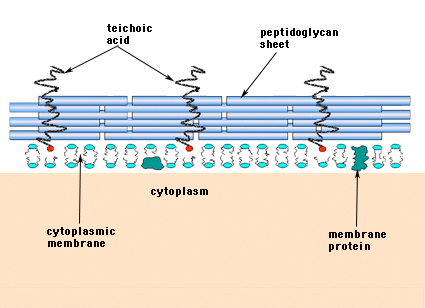
Figure 14. Estrutura da parede celular da bactéria Gram-positiva. A parede é espessa e consiste em muitas camadas de peptidoglicanos intercaladas com ácidos teicóicos que correm perpendicularmente às folhas de peptidoglicanos.
Na bactéria Gram-negativaBacteria (que não retém o violeta cristal), a parede celular é composta por uma única camada depeptidoglicanos circundada por uma estrutura membranosa chamada membrana externa. A membrana externa da bactéria Gram-negativa invariavelmente contém auniquecomponente, lipopolissacarídeo (LPS ou endotoxina), que é tóxico para os animais. Nas bactérias Gram-negativas, a membrana externa é geralmente considerada como parte da parede celular (Figura15).

Figura15. Estrutura da parede celular de Gram-negativos. A parede é relativamente fina e contém muito menos peptidoglicano do que a parede Gram-positiva, além disso, ácidos teicóicos estão ausentes. Entretanto, os wallconsists de células Gram-negativas de uma membrana externa que está fora da camada de peptidoglicano. A membrana externa é ligada à folha de peptidoglicano por um grupo único de moléculas lipoproteicas.
Na Bactéria Gram-positiva, a parede celular é espessa(15-80nanômetros), consistindo de várias camadas de peptidoglicano. No grupo Gram-negativoBactérias, a parede celular é relativamente fina (10 nanômetros) e é composta de uma única camada de peptidoglicano circundada por uma membrana externa.
Estrutura e disposição dos peptidoglicanos em E.coli é representativa de todas as Enterobacteriaceae, assim como de muitas outras bactérias Gram-negativas. A espinha dorsal da glicana é composta por moléculas de N-acetilglucosamina (G) e ácido N-acetilmurâmico (M)ligadas por uma ligação beta 1,4-glicosídeo. O ácido 3-carbono de N-acetilmurâmico (M)é substituído por um grupo de éter lactílico derivado do piruvato. O éter láctico liga a espinha dorsal de glicano a uma cadeia lateral de peptídeo que contém L-alanina, (L-ala), D-glutamato (D-glu), ácido diaminopimélico (DAP), e D-alanina (D-ala). MurNAc é exclusivo das paredes celulares bacterianas, assim como o isD-glu, DAP e D-ala. A subunidade ácido murâmico de E. coli é mostrada emFigure16 abaixo.

Figure 16. A estrutura da subunidade ácido murâmico do peptidoglicano de Escherichia coli. Este é o tipo de mureína encontrada na maioria das bactérias Gram-negativas. Theglycanbackbone é um polímero de repetição de dois aminoácidos, N-acetilglucosamina(G)e ácido N-acetilmurâmico (M). Anexado ao ácido N-acetilmurâmico está o atetrapéptido constituído por L-ala-D-glu-DAP-D-ala. b. Estrutura abreviada da subunidade ácido murâmico. c. Correntes laterais tetrapeptídicas próximas podem estar ligadas umas às outras por uma ligação interpeptídica entre DAP em uma corrente e D-ala na outra. d. A forma polimérica da molécula.
Tiranhas de mureinare montadas no periplasma de cerca de 10 subunidades de ácido murâmico. Onde quer que a sua proximidade o permita, as cadeias de tetrapéptidos que se projectam da espinha dorsal do gliccano podem ser interligadas por uma ligação interpéptida entre um grupo amino livre em DAP e um grupo carboxi livre numa D-ala próxima.A montagem do peptidoglicano no exterior da membrana plasmática é mediada por um grupo de enzimas perplasmáticas, que são transglicosilases, transpeptidases e carboxipeptidases. O mecanismo de ação da penicilina e dos antibióticosbeta-lactâmicos relacionados é o bloqueio da transpeptidase e carboxipeptidaseenzimas durante a sua montagem da parede celular da mureína. Assim, diz-se que os antibióticos thebetalactam “bloqueiam a síntese da parede celular” nas bactérias.
A espinha dorsal do glicano do peptidoglicânmoleculecan é clivada por uma enzima chamada lisozima que está presente no inanimalserum, tecidos e secreções, e no lisossoma fagocitário. A função da lisozima é a de lise das células bacterianas como um agente patogénico constitutivo de defesa contra bactérias. Algumas bactérias Gram-positivas são tolissozimas muito sensíveis e a enzima é bastante activa em baixas concentrações. As lacerações (lacerações) podem ser diluídas 1:40,000 e retêm a capacidade de tolerar as células bacterianas. As bactérias gram-negativas são menos vulneráveis ao ataque da lisozima porque o seu peptidoglicano é protegido pela outermembrana. O local exato da clivagem lisozimal é o beta 1,4 entre o ácido N-acetilmurâmico (M) e a N-acetilglucosamina (G) , de tal forma que a subunidade de ácido lisozimático mostrada na Figura 16(a) é o resultado da ação do peptidoglicano da bactéria lisozima.
Em bactérias Gram-positivas existem arranjos numerousdiferentes de peptidoglicanos. O melhor estudado é a teureína de Staphylococcus aureus mostrada na Figura 17 abaixo. No lugar de DAP(inE. coli) está o ácido diamino, L-lisina (L-lis), e no lugar da ligação interpeptídica (em Gram-negativos) está uma ponte interpeptídica de aminoácidos que liga um grupo de aminoácidos livres na lisina a um grupo freecarboxi em D-ala de uma cadeia lateral tetrapeptídica próxima. Isto permite uma ligação cruzada mais frequente entre as cadeias laterais tetrapeptídicas vizinhas. Em S. aureus, a ponte interpeptídea é um peptideconsistingof 5 moléculas de glicina (chamada ponte de pentaglicina). A montagem da ponte interpeptídica em Gram-positivo mureína é inibida pelos antibióticos beta-lactâmicos da mesma forma que a ligação interpeptídica em Gram-negativo mureína. As bactérias Gram-positivas são bactérias topenicilínticas Gram-negativas mais sensíveis porque o peptidoglicano não está protegido por uma membrana externa e é uma molécula mais abundante. Nas bactérias Gram-positivas, os peptidoglicanos podem variar no aminoácido no lugar de DAP ou L-lis na posição 3 do tetrapeptídeo, e na composição exata da ponte interpeptídea. Pelo menos oito tipos diferentes de peptidoglicanos podem existir em bactérias Gram-positivas.
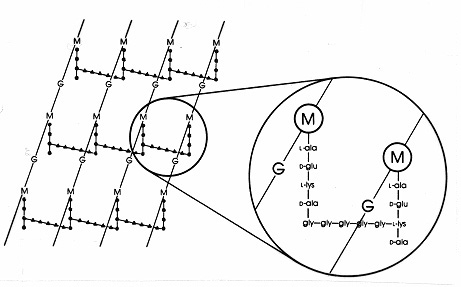
Figure 17. Diagrama esquemático da folha de peptidoglicano de Staphylococcus aureus. G = N-acetil-glucosamina; M = ácido N-acetil-murâmico; L-ala = L-alanina; D-ala = D-alanina; D-glu = ácido D-glutâmico; L-lys = L-lysina. Este é um tipo de mureína encontrada em bactérias Gram-positivas. Comparado ao E. coli peptidoglicano (Figura7) há L-lys no lugar de DAP (ácido diaminopimélico) no tetetrapéptido. O grupo amino livre de L-lys é substituído por um glicinepentapéptido (glicinepentapéptido) que então se torna uma ligação interpeptídica bridgeforminga com um grupo carboxi de D-ala em um tetrapéptido sidechain adjacente. Os peptídeos Gram-positivos diferem de espécie para espécie, principalmente no que diz respeito aos aminoácidos na terceira posição da cadeia tetrapeptídea e na composição de aminoácidos da ponte interpeptídica.
Bacteriamay Gram-negativo contém uma camada única de mureína nas paredes celulares, enquanto as bactérias Gram-positivas são consideradas como tendo várias camadas ou “envoltórios” de peptídeos. Estreitamente associado às camadas de peptidoglicano nas bactérias Gram-positivas está um grupo de moléculas chamadas ácidos teicóicos. Os ácidos teicóicos são polímeros lineares de poliglicerol ou poliribitol substituídos por fosfatos e alguns aminoácidos e açúcares. Os polímeros do ácido teico são ocasionalmente ancorados à membrana plasmática (chamada ácido lipoteico, LTA) aparentemente dirigida para fora em ângulo reto com as camadas de peptidoglicano. As funções do ácido teico não são conhecidas. Eles são essenciais para a viabilidade das bactérias Gram-positivas na natureza. Uma idéia é que eles fornecem um canal de cargas negativas de orientação regular para o lançamento de substâncias com carga positiva através da complicada rede de peptidoglicanos. Outra teoria é que os teicócidos estão de alguma forma envolvidos na regulação e montagem de unidades de ácidos murâmicos no exterior da membrana plasmática. Existem casos, particularmente nos estreptococos, onde os ácidos teicóicos têm sido implicados na aderência das bactérias às superfícies dos tecidos.
capítulo continuado
Página anterior
Deixe uma resposta