Semana del tiburón: ¿Qué comía el Megalodón?
On diciembre 13, 2021 by adminAutor invitado: Jack Cooper, MSc
UoB Graduate / PhD Student, University of Swansea
Como hemos visto hasta ahora, ha habido cambios y actualizaciones en el consenso científico de la asignación taxonómica del Megalodon y su tamaño corporal máximo. Sin embargo, su bocado favorito es algo en lo que la comunidad científica está universalmente de acuerdo. Otra rareza es que es algo que suele representarse con bastante exactitud en la ficción. Permítanme presentarles a un tiburón que comía ballenas, ¡ballenas de verdad!
Muchos de los grandes tiburones macropredatorios actuales son conocidos por comer mamíferos marinos, especialmente nuestro viejo amigo el gran tiburón blanco. Algunas de las fotografías más icónicas de este tiburón incluyen cómo se lanza fuera del agua con su última víctima entre las mandíbulas (Fig. 1). Sin embargo, los tiburones blancos juveniles no comen focas desde el principio. En cambio, empiezan comiendo peces y tiburones más pequeños. Si volvemos a mirar la formación de Gatún en Panamá, encontramos muchos fósiles de tiburones, rayas y otolitos (estructuras del oído interno de los peces), así como una plétora de invertebrados como moluscos.
Algo que no encontramos, sin embargo, son fósiles de mamíferos marinos. Esto parece indicar que las crías de Megalodon también comían peces y tiburones más pequeños en lugar de ir inmediatamente tras los mamíferos marinos. Esto es algo que se llama un cambio ontogenético en la dieta, lo que significa que el tiburón cambia su fuente de alimentación preferida a medida que crece. Al comer alimentos más grandes, y probablemente en gran cantidad, al alcanzar su tamaño adulto, el Megalodon habría podido mantener las exigencias metabólicas de ser tan grande. Podría haber hecho esto comiendo no sólo ballenas, sino casi cualquier cosa que se encontrara.

Dada la cantidad de comida que necesitaba el Megalodon para justificar su enorme tamaño, es casi seguro que se alimentaba de forma oportunista. El registro fósil es nuestra mejor guía aquí. Esto se debe a la presencia de rastros fósiles, que son registros geológicos de la actividad biológica. Esto puede ir desde las huellas de dinosaurios a los arañazos en los huesos que indican que algo mordió al animal al que pertenecía el hueso. Y el Megalodon dejó algunos de estos fósiles de rastreo, proporcionando evidencia clave de sus matanzas.
Se han encontrado varios fósiles pertenecientes a una variedad de cetáceos con notables marcas de mordeduras en ellos. Un ejemplo bien citado proviene de la formación Yorktown en Maryland, donde varios huesos de ballena tenían marcas de mordeduras que habían sido claramente hechas por grandes dientes dentados . De hecho, a veces se encuentran dientes de Megalodón directamente asociados a estos huesos.
Los tiburones pierden y reemplazan los dientes miles de veces a lo largo de su vida, y por lo tanto los tiburones frecuentemente pierden dientes cuando realizan una matanza violenta. Esta es una razón clave por la que los dientes de tiburón son fósiles tan comunes. La presencia de dientes de tiburón justo al lado de un hueso de ballena rayado indica que se estaba alimentando. Pero, algo que los paleontólogos siempre deben considerar cuando se encuentran estos fósiles es: ¿fue realmente una depredación? La búsqueda de comida es un comportamiento común en los tiburones macropredatorios modernos, como los grandes blancos (Fig. 2) y los tiburones tigre. Cuando estos tiburones se encuentran con ballenas muertas flotando en la superficie, es un buffet de todo lo que puedas comer. Lo más importante es que el carroñeo no requiere ningún gasto de energía, algo que puede desperdiciarse en un intento de depredación fallido. Dado que el Megalodon necesitaba ser capaz de mantener su enorme tamaño, no tendría mucho sentido dejar pasar una comida gratis si se encontrara con una ballena muerta. Por lo tanto, por aproximación a cómo se alimentan los tiburones modernos de gran tamaño, la mayoría de nosotros estaríamos dispuestos a apostar que el Megalodon probablemente carroñeara cuando se le daba la oportunidad.

Dicho esto, hay un fósil particularmente notable que implica al Megalodon en un intento de depredación. Y, fundamentalmente, lo sabemos porque su intento fracasó. Un artículo publicado en 2010 describe un fósil parcial del Plioceno perteneciente a un cetáceo de la formación Yorktown en Carolina del Norte (sí, dos formaciones Yorktown). Este fósil también mostraba daños que se asemejaban a marcas de mordeduras, pero, en particular, parecía haber una cubierta ósea tejida sobre estas marcas. Los autores interpretaron esto como un signo de infección que se había producido mientras el animal se recuperaba del traumatismo óseo causado por esas marcas de mordedura. Estas marcas coincidían con una mordedura dentada, lo que sugería que el atacante tenía dientes dentados, al igual que el Megalodon. Por lo tanto, el estudio sugirió que un Megalodon, u otro tiburón de gran tamaño, había atacado al animal del que procedía el fósil, pero la víctima había sobrevivido al intento sólo para morir de una infección por su herida seis semanas después.
Incluso más recientemente, las primeras marcas de mordedura fósiles encontradas en el hemisferio sur atribuidas a Megalodon fueron encontradas en la formación Pisco de Perú. Tal vez estaba destinado a ser, ya que este es el mismo sitio donde Gordon Hubbell encontró su excepcional fósil de Carcharodon (discutido en Q1). El material mordido encontrado aquí incluía restos de cráneo y costillas, lo que sugiere que el tiburón había mordido a su objetivo desde un lado. Lo más significativo es que, por primera vez, los restos fueron identificados como una especie individual: esa especie es Piscobalaena nana, una pequeña ballena con barbas. Al igual que en los otros rastros fósiles, las marcas de mordedura coincidían con los grandes dientes aserrados, un buen argumento para acusar a Megalodon del ataque (Fig. 3). Por lo tanto, parece que el Megalodon también era capaz de atacar a ballenas más pequeñas (ya sea por depredación o por carroñeo), muy en consonancia con las sugerencias de que probablemente también atacaba a delfines, tortugas marinas y otras presas más pequeñas.

Aunque las estrategias de depredación son difíciles de interpretar a partir de los fósiles, los tiburones modernos pueden darnos una idea de cómo cazaba el Megalodon. Su fuerza de mordida habría sido excepcionalmente poderosa para dañar los huesos de las ballenas. Un modelo informático de un cráneo de tiburón blanco ha propuesto que tiene una fuerza de mordida superior a unas 1,8 toneladas, lo que le confiere una de las fuerzas de mordida más fuertes de cualquier animal vivo. Por extensión, este estudio sugiere que la fuerza de mordida de Megalodon era aún mayor, estimada en unos extraordinarios 108.514-182.201 N.
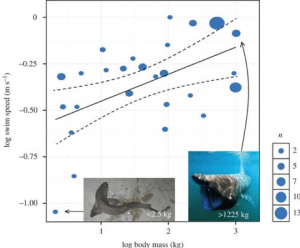
Los científicos también han intentado calcular la velocidad de natación de Megalodon, lo que puede ayudar a darnos una indicación de cómo cazaba. Los tiburones pueden tener una velocidad de ráfaga, que es una sacudida rápida de natación veloz, normalmente utilizada para agarrar la presa. Pero su velocidad habitual para sus nados ocasionales es lo que llamamos velocidad de nado sostenida. Esto varía según la especie, pero un estudio dirigido por David Jacoby combinó datos de 26 especies de 64 estudios anteriores para desarrollar un modelo que predijera una relación de escala entre la velocidad de nado sostenida y el metabolismo y la masa corporal del tiburón. Su modelo, que incorpora el metabolismo, reveló un exponente de escala de 0,173, que se resume en la siguiente ecuación:
Velocidad de nado (ms ¯¹) ∝ Masa (kg)^0,173
Sin embargo, cuando corrigieron los datos en función de la filogenia, descubrieron que la velocidad mínima de nado se escalaba con la masa corporal con un exponente ligeramente inferior de 0,15. La ecuación anterior asume además una constante de 1 para ajustarse a las proporciones, pero los datos revelaron que esta constante era en realidad de 0,266. Por lo tanto, construyeron la ecuación de potencia:
Velocidad de nado (ms ¯¹) = 0,266 Masa (kg) ^0,15
A partir de esto, son capaces de calcular la velocidad de nado sostenida del Megalodon en función de su masa. Por ejemplo, el Megalodon de ~48.000 kg calculado como la masa de un tiburón de 15,9 m en el trabajo de Gottfried revelaría un tiburón nadando continuamente a 1,34 m/s – aproximadamente 4,8 km/h . Aplicando esto a un Megalodón de 52.000 kg se obtiene una velocidad de nado sostenida de 1,36 m/s, es decir, 4,9 km/h, mientras que si vamos al extremo y simulamos un tiburón de 100.000 kg, este modelo daría como resultado una velocidad de nado de 1,50 m/s o 5,4 km/h. Este modelo ha demostrado ser muy eficaz no sólo para calcular la velocidad de nado del Megalodon, sino también para proporcionar un modelo para múltiples especies de tiburones (Fig. 4).
Sin embargo, es la velocidad de ráfaga la que utilizan los tiburones para arrebatar rápidamente sus presas. Los tiburones blancos la utilizan para su «comportamiento de brecha», en el que se impulsan fuera del agua abalanzándose sobre la presa desde abajo para atraparla por sorpresa (Fig. 1) . Pueden hacer esto porque están bien camuflados debido al contraluz. Por lo tanto, dado que el Megalodon probablemente también se camuflaba, y tenía una ecología y una estructura corporal similar a la de los tiburones blancos, esto plantea la pregunta: ¿el Megalodon también era capaz de brincar?
Tal vez. La velocidad de ruptura del tiburón blanco es impulsada por una adaptación fisiológica llamada mesotermia . Esto es algo que recientemente se ha sugerido que el Megalodon también tenía (que se discutirá en detalle en la P5). Los peces mesotérmicos generalmente tienen velocidades de natación sostenidas y de ráfaga más rápidas que los peces ectotérmicos, y esto se ha utilizado para sugerir que el Megalodon puede haber tenido una velocidad de ráfaga de hasta 37,15 km/h.
Esa velocidad increíblemente rápida para un tiburón tan grande bien puede ser suficiente para ser capaz de romper. Además, el tiburón peregrino (Cetorhinus maximus), el segundo tiburón más grande de la actualidad, también ha sido observado haciendo una brecha. Tal vez el gigantesco Megalodón era capaz de saltar fuera del agua mientras se apoderaba de su última comida. Todavía no lo sabemos, pero mi opinión personal y (en cierto modo) profesional es que bien podría haber sido posible.
Con un menú tan amplio para elegir, el Megalodon era un depredador de primer orden, sin depredadores propios conocidos. Esto significa dos cosas importantes. En primer lugar, es probable que tuviera una fuerte competencia por sus presas, algo que puede haber contribuido a su extinción. En segundo lugar, como depredador principal, el Megalodón habría sido de suma importancia para su ecosistema, ya que habría mantenido las poblaciones de sus presas bajo control y, por lo tanto, habría preservado un ecosistema saludable.
Por eso los tiburones de hoy son tan importantes para los entornos marinos. La pérdida de un tiburón depredador de primer orden produce efectos irreversibles en cascada que cambian la comunidad que deja atrás. Así, nuestros ecosistemas marinos son como son en parte porque el Megalodón ya no forma parte de ellos. Y las enormes ballenas de los océanos modernos pueden estar tranquilas como resultado.
- Estrada JA, Rice AN, Natanson LJ & Skomal GB 2006. Uso del análisis isotópico de las vértebras en la reconstrucción de la ecología alimentaria ontogenética en los tiburones blancos. Ecology 87, 829-834.
- Pimiento C, Ehret DJ, MacFadden BJ & Hubbell G 2010. Antigua zona de cría del extinto tiburón gigante Megalodon del Mioceno de Panamá. PLoS One 5, e10552.
- Pimiento C, González-Barba G, Ehret DJ, Hendy AJ, MacFadden BJ & Jaramillo C 2013. Tiburones y rayas (Chondrichthyes, Elasmobranchii) de la formación Gatún del Mioceno tardío de Panamá. J. Paleontol. 87, 755-774.
- Purdy RW 1996. Paleoecología de los tiburones blancos fósiles. En: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 67-78, Academic Press, San Diego.
- Aguilera OA, García L & Cozzuol MA 2008. Tiburones blancos de dientes gigantes e interacción trófica con cetáceos de la Formación Paraguaná del Plioceno del Caribe. Paläontol. Z. 82, 204-208.
- Fallows C, Gallagher A. & Hammerschlag N 2013. Los tiburones blancos (Carcharodon carcharias) hurgan en las ballenas y su papel potencial en la formación de la ecología de un depredador ápice. PLoS One, 8, e60797.
- Dudley SFJ, Anderson-Reade MD, Thompson GS & McMullen PB 2000. Carroñeo simultáneo de un cadáver de ballena por parte de tiburones blancos, Carcharodon carcharias, y tiburones tigre, Galeocerdo cuvier. Fish. Bull. 98, 646-649.
- Kallal RJ, Godfrey SJ & Ortner DJ 2010. Las reacciones óseas en una costilla de cetáceo del Plioceno indican la supervivencia a corto plazo de un evento de depredación. Int. J. Osteoarchaeol. 22, 253-260.
- Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M & Bianucci G 2017. El tiburón gigante extinto Carcharocles megalodon, ¿se dirigía a presas pequeñas? Marcas de mordedura en restos de mamíferos marinos del Mioceno tardío de Perú. Palaeogeogr. Palaeoclimatol. Palaeoecol. 469, 84-91.
- Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, Ferrara TL, Cunningham E, Dean MN & Summers AP 2008. Análisis tridimensional por ordenador de la mecánica de la mandíbula del tiburón blanco: ¿con qué fuerza puede morder un gran blanco? J. Zool. 276, 336-342.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2015. Está la escala de la velocidad de nado en los tiburones impulsada por el metabolismo? Biol. Lett. 11, 20150781.
- Gottfried MD, Compagno LJV & Bowman SC 1996. Tamaño y anatomía del esqueleto del tiburón gigante «megatodo» Carcharodon megalodon. En: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 55-66, Academic Press, San Diego.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2016. Corrección a ‘¿Es la escala de la velocidad de nado en los tiburones impulsada por el metabolismo?’ Biol. Lett. 12, 20160775.
- Watanabe YY, Payne NL, Semmens JM, Fox A & Huveneers C 2019. Estrategias de natación y energética de los tiburones blancos endotérmicos durante la búsqueda de alimento. J. Exp. Biol. 222, jeb185603.
- Cooper JA, Pimiento C, Ferrón HG & Benton MJ (en prensa). Dimensiones del cuerpo del extinto tiburón gigante Otodus megalodon: Una reconstrucción en 2D. Sci. Rep.
- Watanabe YY, Goldman KJ, Caselle JE, Chapman DD & Papastamatiou YP 2015. Los análisis comparativos de datos de seguimiento de animales revelan la importancia ecológica de la endotermia en los peces. Proc. Nat. Acad. Sci. 112, 6104-6109.
- Ferrón HG 2017. La endotermia regional como desencadenante del gigantismo en algunos tiburones macropredatorios extintos. PLoS One, 12, e0185185.
- Johnston EM, Halsey LG, Payne NL, Kock AA, Iosilevskii G, Whelan B &Houghton JD 2018. El poder latente de los tiburones peregrinos revelado por eventos excepcionales de brecha. Biol. Lett. 14, 20180537.
- Myers RA, Baum JK, Shepherd TD, Powers SP & Peterson CH 2007. Efectos en cascada de la pérdida de tiburones depredadores del ápice de un océano costero. Science, 315, 1846-1850.
Editado por Rhys Charles
Deja una respuesta