Glicosilación ligada a N
On diciembre 18, 2021 by admin
La biosíntesis de los glicanos ligados a N ocurre a través de 3 pasos principales:
- Síntesis del oligosacárido precursor ligado a dolicol
- Transferencia en bloque del oligosacárido precursor a la proteína
- Procesamiento del oligosacárido
La síntesis, la transferencia en bloque y el recorte inicial del oligosacárido precursor se producen en el retículo endoplásmico (RE). El posterior procesamiento y modificación de la cadena de oligosacáridos se lleva a cabo en el aparato de Golgi.
La síntesis de las glicoproteínas está así separada espacialmente en diferentes compartimentos celulares. Por lo tanto, el tipo de N-glicano sintetizado depende de su accesibilidad a las diferentes enzimas presentes en estos compartimentos celulares.
Sin embargo, a pesar de la diversidad, todos los N-glicanos se sintetizan a través de una vía común con una estructura central de glicano común.La estructura central del glicano se compone esencialmente de dos residuos de N-acetilglucosamina y tres de manosa. Este glicano central se elabora y modifica posteriormente, dando lugar a una diversa gama de estructuras de N-glicanos.
Síntesis del oligosacárido precursorEditar
El proceso de glicosilación ligado a N comienza con la formación del azúcar GlcNAc ligado a dolicol. El dolicol es una molécula lipídica compuesta por unidades repetidas de isopreno. Esta molécula se encuentra unida a la membrana del RE. Las moléculas de azúcar están unidas al dolicol a través de un enlace de pirofosfato (un fosfato estaba originalmente unido al dolicol, y el segundo fosfato procedía del azúcar nucleótido). A continuación, la cadena de oligosacáridos se extiende mediante la adición de varias moléculas de azúcar de forma escalonada para formar un oligosacárido precursor.
El ensamblaje de este oligosacárido precursor se produce en dos fases: Fase I y II. La fase I tiene lugar en el lado citoplasmático del RE y la fase II tiene lugar en el lado luminal del RE.
La molécula precursora, lista para ser transferida a una proteína, está formada por 2 GlcNAc, 9 manosas y 3 moléculas de glucosa.

|
|
| Fase II | |
es el donante de residuos de manosa (formación : Dol-P + GDP-Man → Dol-P-Man + GDP) y Dol-P-Gluc es el donante de residuos de glucosa (formación : Dol-P + UDP-Glc → Dol-P-Glc + UDP).
|
|
Transferencia del glicano a la proteínaEditar
Una vez formado el oligosacárido precursor, el glicano completado se transfiere al polipéptido naciente en el lumen de la membrana del RE. Esta reacción es impulsada por la energía liberada de la ruptura del enlace pirofosfato entre la molécula de dolicol-glicano.Hay tres condiciones que deben cumplirse antes de que un glicano sea transferido a un polipéptido naciente:
- La asparagina debe estar localizada en una secuencia consenso específica en la estructura primaria (Asn-X-Ser o Asn-X-Thr o en raras ocasiones Asn-X-Cys).
- La asparagina debe estar localizada adecuadamente en la estructura tridimensional de la proteína (los azúcares son moléculas polares y, por lo tanto, deben estar unidos a la asparagina localizada en la superficie de la proteína y no enterrada dentro de la misma)
- La asparagina debe encontrarse en el lado luminal del retículo endoplásmico para que se inicie la glucosilación ligada al N. Los residuos objetivo se encuentran en las proteínas secretoras o en las regiones de la proteína transmembrana que da al lumen.
La oligosacariltransferasa es la enzima responsable del reconocimiento de la secuencia consenso y de la transferencia del glicano precursor a un aceptor polipeptídico que se está traduciendo en el lumen del retículo endoplásmico. La glicosilación ligada al N es, por tanto, un evento cotraduccional
Procesamiento del glicanoEditar

El procesamiento del glicano N se lleva a cabo en el retículo endoplásmico y el cuerpo de Golgi. El recorte inicial de la molécula precursora se produce en el RE y el procesamiento posterior tiene lugar en el Golgi.
Al transferir el glicano completado al polipéptido naciente, se eliminan dos residuos de glucosa de la estructura. Las enzimas conocidas como glicosidasas eliminan algunos residuos de azúcar. Estas enzimas pueden romper los enlaces glicosídicos utilizando una molécula de agua. Estas enzimas son exoglucosidasas, ya que sólo actúan sobre los residuos de monosacáridos situados en el extremo no reductor del glicano. Se cree que este paso inicial de recorte actúa como un paso de control de calidad en el RE para supervisar el plegado de la proteína.
Una vez que la proteína está plegada correctamente, dos residuos de glucosa son eliminados por la glucosidasa I y II. La eliminación del último tercer residuo de glucosa indica que la glicoproteína está lista para el tránsito desde el RE al cis-Golgi. La mannosidasa del RE cataliza la eliminación de esta última glucosa. Sin embargo, si la proteína no está bien plegada, los residuos de glucosa no se eliminan y, por tanto, la glicoproteína no puede salir del retículo endoplásmico. Una proteína chaperona (calnexina/calreticulina) se une a la proteína desdoblada o parcialmente plegada para ayudar al plegado de la proteína.
El siguiente paso implica la adición y eliminación adicional de residuos de azúcar en el cis-Golgi. Estas modificaciones son catalizadas por glicosiltransferasas y glicosidasas respectivamente. En el cis-Golgi, una serie de mannosidasas eliminan algunos o todos los cuatro residuos de manosa en los enlaces α-1,2. Mientras que en la porción medial del Golgi, las glucosiltransferasas añaden residuos de azúcar a la estructura del núcleo del glicano, dando lugar a los tres tipos principales de glicanos: de alta manosa, híbridos y complejos.
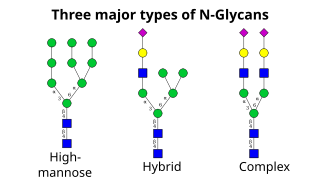
- La alta manosa es, en esencia, sólo dos N-acetilglucosaminas con muchos residuos de manosa, a menudo casi tantos como los que se ven en los oligosacáridos precursores antes de unirse a la proteína.
- Los oligosacáridos complejos se llaman así porque pueden contener casi cualquier número de los otros tipos de sacáridos, incluyendo más de las dos N-acetilglucosaminas originales.
- Los oligosacáridos híbridos contienen un residuo de manosa en un lado de la rama, mientras que en el otro lado una N-acetilglucosamina inicia una rama compleja.
El orden de adición de los azúcares a las cadenas de glicanos en crecimiento está determinado por las especificidades de sustrato de las enzimas y su acceso al sustrato a medida que se mueven por la vía secretora. Así, la organización de esta maquinaria dentro de una célula juega un papel importante en la determinación de los glicanos que se fabrican.
Enzimas del GolgiEditar
Las enzimas del Golgi juegan un papel clave en la determinación de la síntesis de los distintos tipos de glicanos. El orden de acción de las enzimas se refleja en su posición en la pila del Golgi:
| Enzimas | Localización dentro del Golgi |
|---|---|
| Mannosidasa I | cis-Golgi |
| GlcNAc transferasas | medial Golgi |
| Galactosiltransferasa y Sialiltransferasa | trans-Golgi |
En arqueas y procariotasEditar
Se han encontrado vías de biosíntesis de N-glicanos similares en procariotas y arqueas. Sin embargo, en comparación con los eucariotas, la estructura final del glicano en eubacterias y arqueas no parece diferir mucho del precursor inicial fabricado en el retículo endoplásmico. En los eucariotas, el oligosacárido precursor original se modifica ampliamente en su camino hacia la superficie celular.
Deja una respuesta