Fronteras de la Psicología
On septiembre 28, 2021 by adminEl cuerpo calloso (CC) es la comisura primaria que conecta los dos hemisferios cerebrales del cerebro. Por lo tanto, aunque el término comisurotomía no es incorrecto para describir el llamado «cerebro dividido», y es estrictamente correcto para las operaciones quirúrgicas humanas originales en las que se seccionaron todas las comisuras (Gazzaniga et al, 1967), las cirugías posteriores sólo seccionaron el CC, por lo que se utilizó el término «callosotomía», que sigue siendo el más apropiado en esos casos (por ejemplo, Franz et al., 1996, 2000).
Desde una perspectiva evolutiva, el CC es un desarrollo reciente observado sólo en los mamíferos placentarios (Katz et al., 1983; Mihrshahi, 2006). Se cree que la funcionalidad proporcionada por el CC fue precedida, al menos en parte, por la comisura anterior (CA), como se observa en mamíferos no placentarios, como los marsupiales (Heath y Jones, 1971; Ashwell et al., 1996). Aunque algunas vías residuales de la CA todavía están presentes en el cerebro humano, su conectividad y estructura siguen siendo poco estudiadas en la literatura actual y los escasos hallazgos son bastante inconsistentes (Lasco et al., 2002; Patel et al., 2010; Choi et al., 2011). Por ejemplo, los hallazgos basados en imágenes de resonancia magnética (IRM) son contradictorios en cuanto a si el CA es de tamaño similar en mujeres y hombres (Lasco et al., 2002; Patel et al., 2010). En un estudio relativamente reciente en el que se utilizó la técnica in vivo de imágenes con tensor de difusión (DTI), que aprovecha la difusión de las moléculas de agua en los axones, Patel et al. (2010) estudiaron a ocho sujetos y lograron aislar las fibras del CA en cinco de ellos. Entre los hallazgos se encontró, en un sujeto, una gran proporción de fibras en la rama posterior del CA que viajaban bilateralmente hacia las regiones parietales donde la mayoría de ellas terminaban, con un haz más pequeño que terminaba en la circunvolución precentral, y otro haz que entraba en el lóbulo temporal. Estas diferencias individuales se encontraron en todos los participantes, aunque la mayoría de los sujetos mostraron proyecciones bilaterales a las cortezas orbitofrontal, parietal y temporal, a las que el CC no se proyecta claramente. En conjunto, los hallazgos implican que el CA tiene proyecciones tanto primarias como vestigiales a áreas neocorticales a las que el CC no se proyecta, como también han sugerido otros (Di Virgilio et al., 1999; Patel et al., 2010).
En nuestra opinión, el AAA depende en gran medida de las redes neocorticales y de la conectividad con estructuras subcorticales como los ganglios basales y el tálamo (entre otras no discutidas específicamente en nuestra propuesta) (Franz, 2012). Así, cualquier desarrollo evolutivo que involucre al CA podría tener también algún papel en el modelo AAA. Sin embargo, hemos encontrado poca literatura que aborde específicamente este posible vínculo.
El papel del CA se ha estudiado en tareas que probablemente requieran formas de atención, como las que implican atención visual. Por ejemplo, es bien sabido que se han estudiado las poblaciones acallosales en el contexto de tareas que implican integración visual (Gazzaniga et al., 1962, 1965; Sperry, 1968; Levy et al., 1972). Más recientemente, Corballis (1995) trató de abordar la integración visual y la atención en un esfuerzo por explicar las discrepancias anteriores en la investigación sobre el acalso. Corballis propuso un sistema de atención visual dicotómico; una parte (que implica la forma) es automática y está localizada en cada hemisferio, y la otra (que implica el movimiento y la localización) es voluntaria y subcortical. El modelo AAA puede basarse en esta observación y abordar las discrepancias observadas en la literatura. Basándonos en nuestro modelo propuesto, a través de los múltiples niveles de atención existen rutas distintas al CC que podrían mediar la transferencia de información entre hemisferios. Una posibilidad es el circuito de los ganglios basales, que proponemos como potencialmente crítico en la mediación de la atención compartida entre hemisferios (Franz, 2012). Al transeccionar el CC, la transferencia directa de la atención a través de las vías corticales se suprime en gran medida (a menos que, por supuesto, el CA desempeñe un papel de apoyo). Sin embargo, la integración de la información visual basada en la atención podría seguir siendo mediada por vías subcorticales y/o por el CA. De esta manera, el sistema de atención de dos partes observado por Corballis (1995) podría estar presente debido a la mediación basada en la atención de la integración visual a través de la CA (y/o los ganglios basales y los circuitos implicados: Franz, 2012).
Los estudios que implican la visión y la atención no necesitan ser considerados totalmente por separado del sistema que regula la acción motora. Sobre la base de nuestros propios hallazgos (Franz, 2004; Franz y Packman, 2004), varias formas de atención (visual e interna-no visual) están estrechamente vinculadas con la acción motora, y es lógico que una posible clave para entender un posible papel de atención del CA se encuentre en el vínculo entre la acción, la visión y la atención. En un estudio anterior, Franz (2004) hizo que participantes normales dibujaran círculos bimanualmente bajo diferentes manipulaciones de retroalimentación visual, o atención interna a cada mano. El tamaño de los círculos dibujados por una mano que recibía una forma de atención interna se hacía más grande de forma similar a los efectos que se producían con la presencia de retroalimentación visual de esa mano (en relación con la ausencia de atención/retroalimentación visual). Nuestro modelo AAA implica que la atención es un componente clave en la regulación de la producción motora (así como en la selección de la acción motora planificada), y en la medida de las evidencias actuales, no podemos descartar que la CA desempeñe un papel en esa función. Sin embargo, la prueba directa del papel de la CA en cualquier tarea cognitiva no es trivial.
Probar si la CA participa en la función cognitiva es casi imposible en la investigación neuropsicológica dado que no es fácil encontrar participantes con CA localizada (y no conocemos ninguno). Por lo tanto, podría ser mucho más intuitivo evaluar las diferencias neurológicas en la CA normal y correlacionar los datos neurológicos con los datos conductuales. El método de tractografía presentado (que dilucida el CA) puede aplicarse con relativa facilidad a grandes muestras de la población normal, e idealmente, puede combinarse con el rendimiento (basado en tareas realizadas durante sesiones de prueba separadas de la colección de DTI) de esos mismos individuos utilizando tareas cognitivas diseñadas a propósito. En este caso, demostramos que la DTI puede llevarse a cabo y puede producir resultados utilizables en una pequeña muestra de personas de la población neurológicamente normal. La propuesta general sería llevar a cabo métodos similares en una muestra de mayor tamaño, y (como se sugirió anteriormente), correlacionar los resultados (de una medida proxy de la densidad de la CA, por ejemplo) con las variables en una tarea cognitiva.
En la Figura 1 mostramos los resultados del rastreo DTI en los datos obtenidos de 10 participantes femeninas obtenidas de un conjunto de datos DTI público en la Universidad Johns Hopkins (ver Agradecimientos para la fuente completa). La edad media era de 22 años (SD = 2,5). Las imágenes se procesaron utilizando la caja de herramientas de difusión FSL (Smith et al., 2004; Woolrich et al., 2009; Jenkinson et al., 2012) y los procedimientos emplearon los utilizados en estudios anteriores sobre otros tractos de fibras (FDT; Behrens et al., 2003a,b, 2007; Johansen-Berg et al., 2004). Brevemente, primero se concatenaron las imágenes en volúmenes 4d para cada participante y se extrajeron los cráneos utilizando la Herramienta de Extracción Cerebral de FSL (BET; Smith, 2002; Jenkinson et al., 2005). Las imágenes extraídas se corrigieron para tener en cuenta el movimiento menor de la cabeza y las corrientes parásitas generadas por los cambios en la dirección del campo. Los tensores de difusión se ajustaron para generar imágenes que muestran las direcciones de difusión primarias y los niveles de FA en cada vóxel para ser utilizados para los mapas de color durante el enmascaramiento. Se realizó una estimación bayesiana de los parámetros de difusión obtenidos mediante técnicas de muestreo con fibras cruzadas (BEDPOSTX; la «X» implica fibras cruzadas) para generar una distribución del tamaño y la dirección de la difusión teniendo en cuenta dos fibras cruzadas para realizar posteriormente la tractografía a través de los vóxeles de difusión.
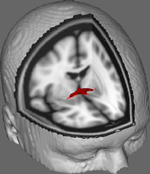
Figura 1. Resultados de rastreo promediados en 10 participantes de control superpuestos en una imagen de IRM ponderada en T1 estándar con vóxeles isotrópicos de 2 mm para referencia anatómica.
Los resultados de rastreo promediados en los 10 participantes de control se superpusieron en una imagen de IRM ponderada en T1 estándar con vóxeles isotrópicos de 2 mm para referencia anatómica. La relación FA media del grupo fue de 0,3146 (SD = 0,0472). Estos resultados de la tractografía revelan, de manera importante, que la estructura de la CA puede ser dilucidada, y por lo tanto tiene potencial para estudios adicionales que correlacionen las puntuaciones de rendimiento en las tareas de comportamiento con los valores de FA utilizando un tamaño de muestra más grande.
En resumen, en la actualidad, existe una escasa literatura, y de la literatura disponible ninguna vincula específicamente la CA a los procesos relacionados con el modelo AAA que es el foco de nuestro artículo objetivo anterior (Franz, 2012). Utilizando un procedimiento similar al propuesto aquí, es posible probar tamaños de muestra más grandes de la población normal, y correlacionar los valores de FA de los resultados del DTI (por ejemplo) con una variable dependiente específica obtenida de una tarea de atención a la acción desarrollada a propósito (cuyos procedimientos son un desarrollo en curso en nuestro laboratorio).
Declaración de conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un potencial conflicto de intereses.
Agradecimientos
Agradecemos al Proyecto Cerebro Humano y al Centro Nacional de Recursos de Investigación (EEUU) y a la Universidad John Hopkins por permitir el uso público de sus datos DTI. También agradecemos el generoso apoyo de la Marsden Fund de la Royal Society de Nueva Zelanda (a Elizabeth A. Franz).
Corballis, M. (1995). Integración visual en el cerebro dividido. Neuropsychologia 33, 937-959.
Pubmed Abstract | Pubmed Full Text
Franz, E. A. (2012). La asignación de la atención al aprendizaje de acciones dirigidas a un objetivo: un marco de neurociencia cognitiva centrado en los ganglios basales. Front. Psychol. 3:535. doi: 10.3389/fpsyg.2012.00535
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heath, C., y Jones, E. (1971). Vías interhemisféricas en ausencia de un cuerpo calloso. An experimental study of commissural connexions in the marsupial phalanger. J. Anat. 109, 253-270.
Pubmed Abstract | Pubmed Full Text
Deja una respuesta