Estructura y función de las células bacterianas
On octubre 10, 2021 by adminEstructura y función de las células bacterianas (página 5)
(Este capítulo tiene 10 páginas)
© Kenneth Todar, PhD
Pared celular
Las paredes celulares de las bacterias merecen una atención especial por varias razones:
1. Son una estructura esencial para la viabilidad, como se ha descrito anteriormente. Son una estructura esencial para la viabilidad, como se ha descrito anteriormente.
2. Están compuestas por componentes únicos que no se encuentran en ningún otro lugar de la naturaleza.
3. Son uno de los sitios más importantes para el ataque de los antibióticos.
4. Proporcionan ligandos para la adherencia y sitios receptores para los fármacos o los virus.
5. Causan síntomas de enfermedad en los animales.
6. Permiten la distinción inmunológica y la variación inmunológica entre las cepas de bacterias.
La mayoría de los procariotas tienen una pared celular rígida.La pared celular es una estructura esencial que protege el protoplasto de la célula del daño mecánico y de la ruptura osmótica o la lisis.Los procariotas suelen vivir en entornos relativamente diluidos, de modo que la acumulación de solutos en el interior del citoplasma de la célula procariota supera ampliamente la concentración total en el entorno exterior. Así, la presión osmótica contra el interior de la membrana plasmática puede ser el equivalente a 10-25atm. Dado que la membrana es una estructura plástica y delicada, debe estar restringida por una pared exterior hecha de un material poroso y rígido que tenga una alta resistencia a la tracción. Este material es la mureína, el componente omnipresente de las paredes celulares bacterianas.
La mureína es un tipo único de peptidoglicano, un polímero de disacáridos (glicano) reticulado por cadenas cortas de aminoácidos (péptido). Existen muchos tipos de peptidoglicanos. Todos los peptidoglicanos bacterianos contienen ácido N-acetilmurámico, que es el componente definitivo de la mureína. Las paredes celulares de las arqueas pueden estar compuestas por proteínas, polisacáridos o moléculas similares al peptidoglicano, pero nunca contienen mureína. Esta característica distingue a las bacterias de las arqueas.
En las bacterias Gram-positivas (las que retienen el colorante violeta de cristal púrpura cuando se someten al procedimiento de tinción de Gram), la pared celular consta de varias capas de peptidoglicano. Perpendicularmente a las láminas de peptidoglicano hay un grupo de moléculas denominadas ácidos teicoicos que son exclusivas de la pared celular de las bacterias Gram positivas (Figura 14). 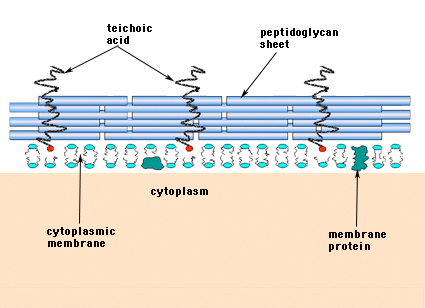
Figura 14. Estructura de la pared celular de las bacterias Gram-positivas. La pared es relativamente gruesa y está formada por muchas capas de peptidoglicano intercaladas con ácidos teicoicos que corren perpendiculares a las láminas de peptidoglicano.
En las bacterias Gram negativas (que no retienen el cristal violeta), la pared celular está compuesta por una sola capa de peptidoglicanos rodeada por una estructura membranosa llamada membrana externa.La membrana externa de las bacterias Gram negativas contiene invariablemente un único componente, el lipopolisacárido (LPS o endotoxina), que es tóxico para los animales. En las bacterias Gram negativas, la membrana externa suele considerarse parte de la pared celular (Figura 15).

Figura 15. Estructura de la pared celular de las bacterias Gram negativas. La pared es relativamente delgada y contiene mucho menos peptidoglicano que la pared de las células Gram positivas; además, los ácidos teicoicos están ausentes. Sin embargo, la pared de las células Gram negativas consiste en una membrana externa que está fuera de la capa de peptidoglicano. La membrana externa está unida a la lámina de peptidoglicano por un grupo único de moléculas de lipoproteínas.
En las bacterias Gram positivas, la pared celular es gruesa (15-80nanómetros) y está formada por varias capas de peptidoglicano. En las bacterias Gram negativas, la pared celular es relativamente fina (10 nanómetros) y está compuesta por una sola capa de peptidoglicano rodeada por una membrana externa.
La estructura y disposición de los peptidoglicanos en E.coli es representativa de todas las Enterobacteriaceae, así como de muchas otras bacterias gramnegativas. La columna vertebral del glicano se compone de moléculas alternas de N-acetilglucosamina (G) y ácido N-acetilmurámico (M) conectadas por un enlace beta 1,4-glicósido. El carbono 3 del ácido N-acetilmurámico (M) está sustituido por un grupo lactílico derivado del piruvato. El éter láctico conecta la columna vertebral del glicano con una cadena lateral peptídica que contiene L-alanina (L-ala), D-glutamato (D-glu), ácido diaminopimélico (DAP) y D-alanina (D-ala). El MurNAc es exclusivo de las paredes celulares bacterianas, al igual que el D-glu, el DAP y la D-ala. La subunidad del ácido murámico de E. coli se muestra en la figura 16.

Figura 16. Estructura de la subunidad de ácido murámico del peptidoglicano de Escherichia coli, que es el tipo de mureína que se encuentra en la mayoría de las bacterias Gram negativas. El núcleo del glicano es un polímero repetido de dos aminoácidos, la N-acetilglucosamina (G) y el ácido N-acetilmurámico (M). Unido al ácido N-acetilmurámico hay un tetrapéptido formado por L-ala-D-glu-DAP-D-ala. b. Estructura abreviada de la subunidad de ácido murámico. c. Las cadenas laterales tetrapéptidas cercanas pueden estar unidas entre sí por un enlace interpéptido entre DAP en una cadena y D-ala en la otra. d. La forma polimérica de la molécula.
Las cadenas de mureína se ensamblan en el periplasma a partir de unas 10 subunidades de ácido murámico. Cuando su proximidad lo permite, las cadenas de tetrapéptidos que se proyectan desde la columna vertebral del glicano pueden reticularse mediante un enlace interpéptido entre un grupo amino libre en el DAP y un grupo carboxi libre en una D-ala cercana.El ensamblaje del peptidoglicano en el exterior de la membrana plasmática está mediado por un grupo de enzimas periplásmicas, que son transglicosilasas, transpeptidasas y carboxipeptidasas. El mecanismo de acción de la penicilina y los antibióticos betalactámicos relacionados consiste en bloquear las enzimas transpeptidasa y carboxipeptidasa durante su ensamblaje de la pared celular de la mureína. Por lo tanto, se dice que los antibióticos betalactámicos «bloquean la síntesis de la pared celular» en las bacterias.
La espina dorsal del glicano de la molécula de peptidoglicano puede ser escindida por una enzima llamada lisozima que está presente en el suero animal, los tejidos y las secreciones, y en el lisosoma fagocítico. La función de la lisozima es lisar las células bacterianas como defensa constitutiva contra los patógenos bacterianos. Algunas bacterias Gram-positivas son muy sensibles a la lisozima y la enzima es bastante activa a bajas concentraciones.Las secreciones lacrimógenas (lágrimas) pueden diluirse 1:40.000 y conservar la capacidad de lisar determinadas células bacterianas. Las bacterias Gram negativas son menos vulnerables al ataque de la lisozima porque su peptidoglicano está protegido por la membrana externa. El lugar exacto de la escisión de la lisozima es el enlace beta 1,4 entre el ácido N-acetilmurámico (M) y la N-acetilglucosamina (G), de modo que la subunidad de ácido murámico que se muestra en la figura 16(a) es el resultado de la acción de la lisozima sobre el peptidoglicano bacteriano.
En las bacterias Gram-positivas existen numerosas disposiciones de péptidos diferentes entre los peptidoglicanos. El más estudiado es el de Staphylococcus aureus, que se muestra en la figura 17. En lugar de la DAP (en E. coli) está el diaminoácido, L-lisina (L-lis), y en lugar del enlace interpéptido (en los Gram-negativos) hay un puente interpéptido de aminoácidos que conecta un grupo amino libre en la lisina con un grupo carboxi libre en la D-ala de una cadena lateral tetrapéptida cercana. Esta disposición permite, aparentemente, una unión cruzada más frecuente entre las cadenas laterales tetrapéptidas cercanas. En S. aureus, el puente interpéptido es un péptido formado por 5 moléculas de glicina (llamado puente de pentaglicina). El ensamblaje del puente interpéptido en la mureína Gram-positiva es inhibido por los antibióticos betalactámicos de la misma manera que el enlace interpéptido en la mureína Gram-negativa. Las bacterias Gram-positivas son más sensibles a los antibióticos que las Gram-negativas porque el peptidoglicano no está protegido por una membrana externa y es una molécula más abundante. En las bacterias Gram-positivas, los peptidoglicanos pueden variar en el aminoácido que sustituye al DAP o al L-lys en la posición 3 del tetrapéptido, y en la composición exacta del puente interpéptido. Al menos ocho tipos diferentes de peptidoglicanos existen en las bacterias Gram-positivas.
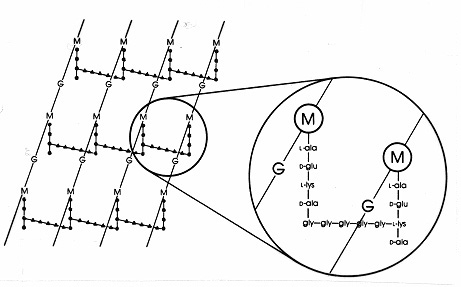
Figura 17. Diagrama esquemático de la hoja de peptidoglicano de Staphylococcus aureus. G =N-acetil-glucosamina; M = ácido N-acetil-murámico; L-ala = L-alanina; D-ala = D-alanina; D-glu= ácido D-glutámico; L-lys = L-lisina. Este es un tipo de mureína que se encuentra en las bacterias Gram-positivas. En comparación con el peptidoglicano de E. coli (Figura 7), hay L-lis en lugar de DAP (ácido diaminopimélico) en el tetrapéptido. El grupo amino libre de L-lis se sustituye por un glicinapéptido (gly-gly-gly-) que se convierte en un puente interpéptido que forma un enlace con un grupo carboxi de D-ala en una cadena lateral tetrapéptida adyacente. Los peptidoglicanos grampositivos difieren de una especie a otra, principalmente en lo que respecta a los aminoácidos en la tercera posición de la cadena de tetrapéptidos y en la composición de aminoácidos del puente interpéptido.
Las bacterias gramnegativas pueden contener una sola capa monomolecular de mureína en sus paredes celulares, mientras que se cree que las bacterias grampositivas tienen varias capas o «envolturas» de peptidoglicano. Las capas de peptidoglicano de las bacterias Gram positivas están estrechamente asociadas a un grupo de moléculas denominadas ácidos teicoicos. Los ácidos teicoicos son polímeros lineales de poliglicerol o polirribitol sustituidos por fosfatos y algunos aminoácidos y azúcares. Los polímeros de ácido teicoico están ocasionalmente anclados a la membrana plasmática (denominados ácido lipoteico, LTA) aparentemente dirigidos hacia el exterior en ángulo recto con las capas de peptidoglicano. No se conocen las funciones del ácido teicoico. Son esenciales para la viabilidad de las bacterias Gram-positivas en la naturaleza. Una de las ideas es que proporcionan un canal de cargas negativas orientadas de forma regular para el paso de sustancias cargadas positivamente a través de la complicada red de peptidoglicano. Otra teoría es que los ácidos teicoicos están de alguna manera implicados en la regulación y el ensamblaje de las subunidades de ácidos murámicos en el exterior de la membrana plasmática. Hay casos, particularmente en los estreptococos, en los que los ácidos teicoicos han estado implicados en la adherencia de las bacterias a las superficies de los tejidos.
continuación del capítulo
Página anterior
Deja una respuesta