Struktur und Funktion von Bakterienzellen
On Oktober 10, 2021 by adminStruktur und Funktion von Bakterienzellen (Seite 5)
(Dieses Kapitel hat 10 Seiten)
© Kenneth Todar, PhD
Zellwand
Die Zellwände von Bakterien verdienen aus mehreren Gründen besondere Aufmerksamkeit:
1. Sie sind eine wesentliche Struktur für die Lebensfähigkeit, wie oben beschrieben.
2. Sie bestehen aus einzigartigen Komponenten, die nirgendwo sonst in der Natur zu finden sind.
3. Sie sind einer der wichtigsten Angriffspunkte für Antibiotika.
4. Sie stellen Liganden für die Adhärenz und Rezeptorstellen für Medikamente oder Viren bereit.
5. Sie verursachen Krankheitssymptome bei Tieren.
6. Sie sorgen für die immunologische Unterscheidung und immunologische Variation zwischen Bakterienstämmen.
Die meisten Prokaryonten haben eine starre Zellwand. Die Zellwand ist eine wesentliche Struktur, die den Zellprotoplasten vor mechanischer Beschädigung und vor osmotischer Ruptur oder Lyse schützt.Prokaryonten leben in der Regel in einem relativ verdünnten Milieu, so dass die Anhäufung von gelösten Stoffen im Zytoplasma der prokaryontischen Zelle die Gesamtkonzentration in der äußeren Umgebung deutlich übersteigt. So kann der osmotische Druck an der Innenseite der Plasmamembran 10-25atm betragen. Da es sich bei der Membran um eine empfindliche, plastische Struktur handelt, muss sie durch eine Außenwand aus porösem, starrem Material mit hoher Zugfestigkeit zusammengehalten werden. Ein solches Material ist Murein, der allgegenwärtige Bestandteil der bakteriellen Zellwände.
Murein ist ein einzigartiger Typ von Peptidoglykan, ein Polymer aus Disacchariden (Glykan), das durch kurze Ketten von Aminosäuren (Peptid) vernetzt ist. Es gibt viele Arten von Peptidoglykanen. Alle bakteriellen Peptidoglykane enthalten N-Acetylmuraminsäure, die der bestimmende Bestandteil von Murein ist. Die Zellwände von Archaea können aus Proteinen, Polysacchariden oder peptidoglykanähnlichen Molekülen bestehen, enthalten aber niemals Murein. Dieses Merkmal unterscheidet die Bakterien von den Archaeen.
Bei den Gram-positiven Bakterien (die den violetten Kristallviolett-Farbstoff behalten, wenn sie dem Gram-Färbeverfahren unterzogen werden) besteht die Zellwand aus mehreren Schichten Peptidoglykan. Senkrecht zu den Peptidoglykanschichten verläuft eine Gruppe von Molekülen, die Teichoinsäuren genannt werden und nur in der Gram-positiven Zellwand vorkommen (Abbildung 14). 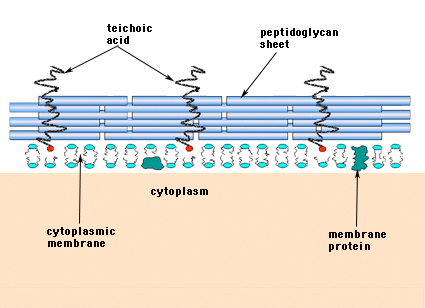
Abbildung 14. Struktur der Gram-positiven Bakterienzellwand. Die Wand ist relativ dick und besteht aus vielen Schichten von Peptidoglykanen, die mit Teichosäuren durchsetzt sind, die senkrecht zu den Peptidoglykanschichten verlaufen.
Bei gramnegativen Bakterien (die das Kristallviolett nicht zurückhalten) besteht die Zellwand aus einer einzigen Schicht von Peptidoglykanen, die von einer Membranstruktur umgeben ist, die als äußere Membran bezeichnet wird.
Die äußere Membran gramnegativer Bakterien enthält ausnahmslos eine einzige Komponente, das Lipopolysaccharid (LPS oder Endotoxin), das für Tiere giftig ist. Bei gramnegativen Bakterien wird die äußere Membran gewöhnlich als Teil der Zellwand betrachtet (Abbildung 15).

Abbildung 15. Struktur der gramnegativen Zellwand. Die Wand ist relativ dünn und enthält viel weniger Peptidoglykan als die Gram-positive Wand, und es fehlen die Teichosäuren. Die gramnegative Zellwand besteht jedoch aus einer äußeren Membran, die sich außerhalb der Peptidoglykanschicht befindet. Die äußere Membran ist durch eine besondere Gruppe von Lipoproteinmolekülen mit der Peptidoglykanschicht verbunden.
Bei den grampositiven Bakterien ist die Zellwand dick (15-80 Nanometer) und besteht aus mehreren Peptidoglykanschichten. Bei den Gram-negativen Bakterien ist die Zellwand relativ dünn (10 Nanometer) und besteht aus einer einzigen Schicht Peptidoglykanen, die von einer äußeren Membran umgeben ist.
Die Struktur und Anordnung der Peptidoglykane in E. coli ist repräsentativ für alle Enterobacteriaceae sowie für viele andere gramnegative Bakterien. Das Glykan-Grundgerüst besteht aus alternierenden Molekülen von N-Acetylglucosamin (G) und N-Acetylmuraminsäure (M), die durch eine Beta-1,4-Glykosidbindung verbunden sind. Der 3-Kohlenstoff der N-Acetylmuraminsäure (M) ist durch eine von Pyruvat abgeleitete Lactylethergruppe substituiert. Der Lactylether verbindet das Glykan-Grundgerüst mit einer Peptid-Seitenkette, die L-Alanin (L-Ala), D-Glutamat (D-Glu), Diaminopimelinsäure (DAP) und D-Alanin (D-Ala) enthält. MurNAc kommt nur in bakteriellen Zellwänden vor, ebenso wie D-Glu, DAP und D-Ala. Die Muraminsäure-Untereinheit von E. coli ist in der folgenden Abbildung 16 dargestellt.

Abbildung 16. Die Struktur der Muraminsäure-Untereinheit des Peptidoglykans von Escherichia coli, dem Mureintyp, der in den meisten gramnegativen Bakterien vorkommt. DasGlykan-Grundgerüst ist ein sich wiederholendes Polymer aus zwei Aminozuckern, N-Acetylglucosamin (G) und N-Acetylmuraminsäure (M). An die N-Acetylmuraminsäure ist ein Tetrapeptid gebunden, das aus L-ala-D-glu-DAP-D-ala besteht. b. Gekürzte Struktur der Muraminsäure-Untereinheit. c. Nahegelegene Tetrapeptid-Seitenketten können durch eine Interpeptidbindung zwischen DAP auf der einen und D-Ala auf der anderen Seite miteinander verbunden sein. d. Die polymere Form des Moleküls.
Mureinstränge werden im Periplasma aus etwa 10 Muraminsäure-Untereinheiten zusammengesetzt. Wo immer es ihre Nähe zulässt, können die Tetrapeptidketten, die aus dem Glykangerüst herausragen, durch eine Interpeptidbindung zwischen einer freien Aminogruppe auf DAP und einer freien Carboxygruppe auf einem benachbarten D-Ala vernetzt werden.Der Aufbau des Peptidoglycans an der Außenseite der Plasmamembran wird durch eine Gruppe von periplasmatischen Enzymen vermittelt, bei denen es sich um Transglycosylasen, Transpeptidasen und Carboxypeptidasen handelt. Der Wirkungsmechanismus von Penicillin und verwandten Beta-Lactam-Antibiotika besteht darin, die Transpeptidase- und Carboxypeptidase-Enzyme beim Aufbau der Murein-Zellwand zu blockieren. Daher sagt man, dass die Betalaktam-Antibiotika die Zellwandsynthese in den Bakterien „blockieren“.
Das Glykan-Grundgerüst des Peptidoglykan-Moleküls kann durch ein Enzym namens Lysozym gespalten werden, das im Tierserum, in Geweben und Sekreten sowie im phagozytischen Lysosom vorhanden ist. Die Funktion von Lysozym ist die Lyse von Bakterienzellen als konstitutiver Schutz vor bakteriellen Krankheitserregern. Einige grampositive Bakterien reagieren sehr empfindlich auf Lysozym, und das Enzym ist schon bei niedrigen Konzentrationen recht aktiv. Tränenflüssigkeit kann im Verhältnis 1:40.000 verdünnt werden und behält die Fähigkeit, bestimmte Bakterienzellen zu lysieren. Gramnegative Bakterien sind weniger anfällig für den Angriff von Lysozym, da ihr Peptidoglykan durch die Außenmembran abgeschirmt ist. Der genaue Ort der lysozymalen Spaltung ist die Beta-1,4-Bindung zwischen N-Acetylmuraminsäure (M) und N-Acetylglucosamin (G), so dass die in Abbildung 16(a) gezeigte Uraminsäure-Untereinheit das Ergebnis der Einwirkung von Lysozym auf bakterielles Peptidoglykan ist.
In Gram-positivenBakterien gibt es zahlreiche unterschiedliche Peptidanordnungen unter den Peptidoglykanen. Das am besten untersuchte ist das in Abbildung 17 gezeigte Theurein von Staphylococcus aureus. Anstelle von DAP (bei E. coli) befindet sich die Diamino-Säure L-Lysin (L-Lys), und anstelle der Interpeptidbindung (bei Gram-negativen) befindet sich eine Interpeptidbrücke aus Aminosäuren, die eine freie Aminogruppe an Lysin mit einer freien Carboxygruppe an D-Ala einer nahe gelegenen Tetrapeptid-Seitenkette verbindet. Diese Anordnung ermöglicht offenbar häufigere Kreuzbindungen zwischen benachbarten Tetrapeptid-Seitenketten. Bei S. aureus ist die Interpeptidbrücke ein Peptid, das aus 5 Glycinmolekülen besteht (sogenannte Pentaglycinbrücke). Der Aufbau der Interpeptidbrücke in Gram-positivem Murein wird durch die Beta-Lactam-Antibiotika auf die gleiche Weise gehemmt wie die Interpeptidbindung in Gram-negativem Murein. Gram-positive Bakterien sind empfindlicher gegenüber Penicillin als Gram-negative Bakterien, da das Peptidoglykan nicht durch eine äußere Membran geschützt ist und es sich um ein häufigeres Molekül handelt. Bei Gram-positiven Bakterien können sich die Peptidoglykane in der Aminosäure anstelle von DAP oder L-Lys in Position 3 des Tetrapeptids und in der genauen Zusammensetzung der Interpeptidbrücke unterscheiden. Mindestens acht verschiedene Typen von Peptidoglykanen kommen in grampositiven Bakterien vor.
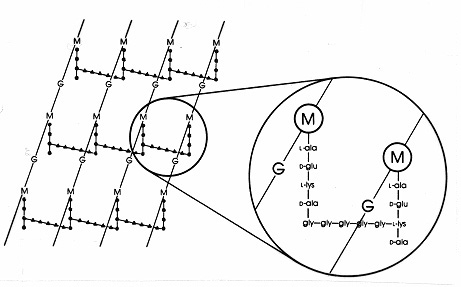
Abbildung 17. Schematische Darstellung der Peptidoglykanschicht von Staphylococcus aureus. G = N-Acetyl-Glucosamin; M = N-Acetyl-Muraminsäure; L-Ala = L-Alanin; D-Ala = D-Alanin; D-Glu = D-Glutaminsäure; L-Lys = L-Lysin. Dies ist ein Typ von Murein, der in Gram-positiven Bakterien vorkommt. Im Vergleich zum Peptidoglykan von E. coli (Abbildung 7) befindet sich L-Lys anstelle von DAP (Diaminopimelinsäure) im Tetrapeptid, wobei die freie Aminogruppe von L-Lys durch ein Glycinpentapeptid (Gly-gly-gly-gly-gly-) ersetzt wird, das dann eine Interpeptidbrücke bildet, die eine Verbindung mit einer Carboxygruppe von D-Ala in einer benachbarten Tetrapeptid-Seitenkette eingeht. Gram-positive Peptidoglykane unterscheiden sich von Spezies zu Spezies, vor allem hinsichtlich der Aminosäuren in der dritten Position der Tetrapeptidkette und der Aminosäurezusammensetzung der Interpeptidbrücke.
Gramm-negative Bakterien können eine einzige monomolekulare Mureinschicht in ihren Zellwänden enthalten, während man davon ausgeht, dass Gram-positive Bakterien mehrere Schichten oder „Hüllen“ aus Peptidoglykan haben. Eng verbunden mit den Peptidoglykanschichten der grampositiven Bakterien ist eine Gruppe von Molekülen, die Teichoinsäuren. Teichoinsäuren sind lineare Polymere aus Polyglycerin oder Polyribitol, die mit Phosphaten und einigen Aminosäuren und Zuckern substituiert sind. Die Teichoinsäure-Polymere sind gelegentlich an der Plasmamembran verankert (Lipoteichoinsäure, LTA), die scheinbar rechtwinklig zu den Peptidoglykanschichten nach außen gerichtet ist. Die Funktionen der Teichoinsäure sind nicht bekannt. In der Natur sind sie für die Lebensfähigkeit von Gram-positiven Bakterien unerlässlich. Eine Idee ist, dass sie einen Kanal mit regelmäßig ausgerichteten negativen Ladungen bilden, um positiv geladene Substanzen durch das komplizierte Peptidoglykan-Netzwerk zu schleusen. Eine andere Theorie besagt, dass Teichoinsäuren in irgendeiner Weise an der Regulierung und dem Zusammenbau von Muraminsäure-Untereinheiten an der Außenseite der Plasmamembran beteiligt sind. Es gibt Fälle, insbesondere bei den Streptokokken, in denen Teichoinsäuren an der Anhaftung der Bakterien an Gewebeoberflächen beteiligt sind.
Kapitel Fortsetzung
Vorige Seite
Schreibe einen Kommentar