Nucleus olivaris superior
On November 12, 2021 by admin| Gehirn: Superior olivary complex | ||
|---|---|---|
 |
||
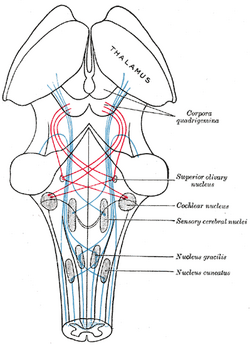
| Schema, das den Verlauf der Fasern des Lemniskus zeigt; medialer Lemniskus in blau, lateraler in rot. (Der Nucleus olivaris superior ist in der Mitte rechts markiert.) | ||
| ] | ||
| Lateinisch | Nucleus olivaris superior | |
| Gray’s | Subjekt #187 787 | |
| Teil von | ||
| Bestandteile | ||
| Arterie | ||
| Vein | ||
| BrainInfo/UW | {{BrainInfoType}}-565 | |
| MeSH | ||
Der Superior Olivary Complex (oder SOC oder Superior Olive) ist eine kleine Masse grauer Substanz, die sich an der dorsalen Oberfläche des lateralen Teils des Trapezkörpers befindet. Beim Menschen ist er klein, bei bestimmten Tieren jedoch gut entwickelt. Er weist eine ähnliche Struktur wie der Nucleus olivaris inferior auf und liegt unmittelbar über diesem. Einige der Fasern des Trapezkörpers enden, indem sie sich um die Zellen dieses Kerns herum verzweigen, während andere von diesen Zellen ausgehen.
Physiologie
Der Nucleus olivaris superior spielt eine Reihe von Rollen beim Hören. Der Nucleus olivaris medialis superior (MSO) ist ein spezialisierter Kern, von dem man annimmt, dass er die Zeitdifferenz der Ankunft von Tönen zwischen den Ohren (die interaurale Zeitdifferenz oder ITD) misst. Die ITD ist ein wichtiger Anhaltspunkt für die Bestimmung des Azimuts von tieffrequenten Tönen, d. h. für deren Lokalisierung in der Azimutalebene, d. h. ihren Grad nach links oder rechts.
Es wird vermutet, dass die laterale Superior-Olive (LSO) an der Messung des Pegelunterschieds von Tönen zwischen den Ohren beteiligt ist (der interaurale Pegelunterschied oder ILD). Die ILD ist ein zweiter wichtiger Anhaltspunkt bei der Bestimmung des Azimuts hochfrequenter Töne.
Beziehung zum auditorischen System
Der Komplex der superioren Olive befindet sich im Allgemeinen im Pons, erstreckt sich aber beim Menschen von der rostralen Medulla bis zum Mittelpons und erhält Projektionen vorwiegend vom anteroventralen Cochlea-Kern über die ventrale akustische Stria, obwohl auch der posteroventrale Kern über die intermediäre akustische Stria dorthin projiziert. An dieser Stelle finden die ersten großen binauralen Interaktionen statt.
Primäre Kerne
Der Komplex der superioren Olivaria unterteilt sich in drei primäre Kerne, den MSO, LSO und den medialen Kern des Trapezkörpers, sowie mehrere kleinere perioliväre Kerne. Diese drei Kerne sind die am besten untersuchten und daher auch am besten verstanden. In der Regel wird davon ausgegangen, dass sie den aufsteigenden azimutalen Lokalisierungsweg bilden.
Mediale Superior Olive (MSO)
Man nimmt an, dass die mediale Superior Olive dabei hilft, den Azimut eines Schalls zu lokalisieren, d. h. den Winkel nach links oder rechts, in dem sich die Schallquelle befindet. Auf den ersten Blick könnte man meinen, dass dieser Kern auch vertikale Informationen enthält, aber das ist nicht der Fall; die im MSO verarbeiteten Informationen sind auf die horizontale Ebene beschränkt. Die fusiformen Zellen projizieren nicht in die Ebene des Pons und kommen erst im Colliculus inferior ins Spiel. Es sind nur horizontale Daten vorhanden, die jedoch von zwei verschiedenen Ohrquellen stammen, was die Lokalisierung des Schalls auf der Azimutachse erleichtert. Dies geschieht, indem die obere Olive die Zeitunterschiede zwischen zwei Ohrsignalen misst, die denselben Stimulus aufnehmen. Die Reise um den Kopf dauert etwa 700 μs, und man geht davon aus, dass die mediale Superior Olive in der Lage ist, dies zu erkennen. Tatsächlich wird beobachtet, dass Menschen interaurale Unterschiede bis zu 10 Mikrosekunden wahrnehmen können. Der Kern ist tonotopisch organisiert, aber die azimutale rezeptive Feldprojektion ist „höchstwahrscheinlich eine komplexe, nichtlineare Karte.“
Die Projektionen der medialen superioren Olive enden dicht im ipsilateralen zentralen Nucleus des Colliculus inferior. Die meisten dieser Axone gelten als „rund“ oder als Typ R. Diese R-Axone sind meist glutamatergisch, enthalten runde synaptische Vesikel und bilden asymmetrische synaptische Verbindungen.
- Dies ist der größte der Kerne und enthält beim Menschen etwa 15.500 Neuronen.
- Jeder MSO erhält niederfrequente bilaterale Eingänge von den rechten und linken AVCNs.
- Der Ausgang geht zum ipsilateralen lateralen Lemniscus und schließlich zum inferioren Colliculus.
- Der MSO reagiert besser auf binaurale Reize.
- Seine Hauptfunktion besteht in der Erkennung von Unterschieden in der Ankunftszeit von Geräuschen an den beiden Ohren, was Teil des Lokalisierungsprozesses (ITD) ist.
Laterale Superior-Olive (LSO)
Diese Olive hat ähnliche Funktionen wie die mediale Superior-Olive, nutzt aber die Intensität zur Lokalisierung der Schallquelle. Dies ist der Teil des Hirnstamms, der den lauteren Schall aus dem linken Ohr als auf der linken Seite befindlich kennzeichnet. Die laterale Olive erhält Input von beiden Cochlea-Kernen, von den ipsilateralen sphärischen Buschzellen des AVCN und den kontralateralen globulären Buschzellen des AVCN über die ipsilaterale MNTB. Die MNTB-Hauptzellen sind glyzinerge Zellen, die eine Quelle der Hemmung für das LSO darstellen und die Grundlage für die ILD-Empfindlichkeit bilden. Die Projektionen aus beiden Cochlea-Kernen sind in erster Linie hochfrequent, und diese Frequenzen werden dann von der Mehrheit der LSO-Neuronen repräsentiert (>2/3 über 2-3 kHz bei der Katze). Zusätzliche Eingänge stammen aus dem ipsilateralen LNTB (glyzinerge Signale, siehe unten), die hemmende Informationen aus dem ipsilateralen Cochlea-Kern liefern. Ein weiterer möglicher inhibitorischer Input stammt von ipsilateralen nicht-sphärischen Zellen des AVCN. Diese Zellen sind entweder kugelförmig-buschig oder multipolar (stellat). Jeder dieser beiden Eingänge könnte die Grundlage für die ipsilaterale Hemmung bilden, die in den Reaktionskarten zu sehen ist, die die primäre Erregung flankieren und die Frequenzabstimmung der Einheit schärfen.
Das LSO projiziert bilateral zum zentralen Kern des Colliculus inferior (ICC). Ipsilaterale Projektionen sind primär hemmend (glyzinerge) und kontralaterale erregend. Weitere Projektionsziele sind die dorsalen und ventralen Kerne des lateralen Lemniskus (DNLL & VNLL). Die GABA-ergen Projektionen aus dem DNLL bilden die primäre GABA-Quelle im auditorischen Hirnstamm und projizieren bilateral zum ICC und zum kontralateralen DNLL. Diese konvergierenden exzitatorischen und inhibitorischen Verbindungen können dazu beitragen, die Pegelabhängigkeit der ILD-Empfindlichkeit im ICC im Vergleich zum LSO zu verringern.
Zusätzliche Projektionen bilden das Lateral Olivocochlear Bundle (LOC), das die inneren Haarzellen der Cochlea innerviert. Es wird angenommen, dass diese Projektionen eine lange Zeitkonstante haben und dazu dienen, den von jedem Ohr erkannten Schallpegel zu normalisieren, um die Schalllokalisierung zu unterstützen. Es gibt beträchtliche Unterschiede zwischen den Arten: Bei Nagetieren sind die LOC-Projektionsneuronen innerhalb des LSO verteilt, während sie bei Raubtieren (z. B. Katzen) den LSO umgeben.
Medialer Nukleus des Trapezkörpers (MNTB)
- Der MNTB ist der kleinste Teil des SOC; das Vorhandensein des MNTB im menschlichen Hirnstamm ist fraglich.
- Jedes Neuron erhält nur hochfrequenten Input über einen Calyx aus dem kontralateralen AVCN, und sein Output projiziert zum ipsilateralen LSO.
- Es gibt zwei Reaktionstypen: einen „Chopper-Typ“, der den Spindelzellen in der AVCN ähnelt, und einen primären Typ, der denen der Buschzellen in der AVCN ähnelt.
Perioliväre Kerne
Der SOC besteht je nach Forscher aus sechs bis neun periolivären Kernen, die in der Regel nach ihrer Lage in Bezug auf die primären Kerne benannt sind. Diese Kerne umgeben jeden der primären Kerne und tragen sowohl zum aufsteigenden als auch zum absteigenden Hörsystem bei. Diese Kerne bilden auch die Quelle des olivocochleären Bündels, das die Cochlea innerviert. Beim Meerschweinchen sind die aufsteigenden Projektionen zu den Colliculi inferior hauptsächlich ipsilateral (>80 %), wobei die größte Einzelquelle aus dem SPON stammt. Auch die ventralen Kerne (RPO, VMPO, AVPO, & VNTB) sind fast vollständig ipsilateral, während die übrigen Kerne bilateral projizieren.
| Name | Katze | Raubtier | Ratte | Maus |
|---|---|---|---|---|
| LSO | X | X | X | X |
| MSO | X | X | X | X |
| MNTB | X | X | X | X |
| LNTB | X | X | „LVPO“ | X |
| ALPO | X | X | ||
| PVPO | X | X | ||
| PPO | X | X | „CPO“ | |
| VLPO | X | |||
| DPO | X | X | X | |
| DLPO | X | X | ||
| VTB | X | X | „MVPO“ | X |
| AVPO | X | |||
| VMPO | X | X | ||
| RPO | X | X | ||
| SPN | „DMPO“ | X | X | X |
Ventraler Nukleus des Trapezkörpers (VNTB)
- Der VNTB ist ein kleiner Kern, der sich lateral zum MNTB befindet, und ventral zum MSO.
- Dieser Kern besteht aus einer heterogenen Zellpopulation, die in viele Hörkerne projiziert und das mediale olivocochleäre Bündel (MOC) bildet, das die äußeren Haarzellen der Cochlea innerviert. Diese Zellen enthalten elektromotile Fasern und wirken als mechanische Verstärker/Dämpfer in der Cochlea.
- Der Kern projiziert in beide IC, wobei keine Zellen bilateral projizieren.
Lateral Nucleus of the Trapezoid Body (LNTB)
- Lokalisiert ventral zum LSO
- AVCN kugelförmige buschige Zellen projizieren Kollaterale bilateral, und globuläre buschige Zellen projizieren Kollaterale ipsilateral zu LNTB-Neuronen.
- Die Zellen sind immunreaktiv für Glycin und werden nach Injektion von tritiiertem Glycin in den LSO retrograd markiert.
- Der Kern projiziert sowohl in den IC, wobei nur wenige Zellen bilateral projizieren, als auch in den ipsilateralen LSO.
- Große multipolare Zellen projizieren sowohl bei der Katze als auch beim Meerschweinchen in den Cochlea-Kern, aber nicht in den IC.
- Die Eingänge erfolgen oft über die Endbläschen, was zu einer sehr schnellen Signaltransduktion führt.
Superiorer periolivärer Nukleus (SPON) (Dorsomedialer periolivärer Nukleus (DMPO))
- Direkt dorsal des MNTB gelegen
- Bei der Ratte ist der SPON ein homogener GABA-erger Nukleus. Diese tonotopisch organisierten Neuronen erhalten exzitatorische Eingänge von Oktopus und multipolaren Zellen im kontralateralen ventralen Cochlea-Kern, einen glyzinergen (hemmenden) Eingang vom ipsilateralen MNTB, einen unbekannten GABAergen (hemmenden) Eingang und projizieren zum ipsilateralen ICC. Die meisten Neuronen reagieren nur auf den Offset eines Reizes, können auf AM-Reize bis zu 200 Hz phasenstarr reagieren und bilden möglicherweise die Grundlage für die ICC-Dauerselektivität. Bemerkenswert ist, dass die SPON-Neuronen keine absteigenden Eingänge vom ICC erhalten, und sie projizieren nicht in die Cochlea oder den Cochlear Nucleus, wie es viele perioliväre Kerne tun.
- Beim Meerschweinchen projizieren runde bis ovale multipolare Zellen in beide IC, wobei viele Zellen bilateral projizieren. Die länglicheren Zellen, die in den Nucleus cochlearis projizieren, projizieren nicht in den ICC. Es scheint zwei Populationen von Zellen zu geben, eine, die ipsilateral projiziert, und eine, die bilateral projiziert.
- Die meisten Informationen stammten von SPON bei Nagetieren, da der Kern bei diesen Spezies sehr groß ist, und es wurden nur sehr wenige Studien an DMPO bei Katzen durchgeführt, von denen keine umfangreich war.
Dorsaler periolivarer Nukleus (DPO)
- Liegt dorsal und medial zum LSO
- Enthält sowohl EE (von beiden Ohren erregt) als auch E0 (nur vom kontralateralen Ohr erregt) Einheiten.
- Neuronen sind tonotopisch organisiert und hochfrequent.
- Können zusammen mit dem DLPO zu einem einzigen Kern gehören
- Der Kern projiziert in beide IC, wobei keine Zellen bilateral projizieren.
Dorsolateraler periolivarer Nukleus (DLPO)
- Liegt dorsal und lateral des LSO
- Enthält sowohl EE (von beiden Ohren erregt) als auch E0 (nur vom kontralateralen Ohr erregt) Einheiten.
- Neuronen sind tonotopisch organisiert und niederfrequent
- können zusammen mit dem DPO zu einem einzigen Kern gehören
- Der Kern projiziert in beide IC, wobei wenige Zellen bilateral projizieren.
Ventrolateraler periolivarer Kern (VLPO)
- Befindet sich ventral und innerhalb des ventralen Hillus des LSO
- Enthält sowohl EI- (erregt durch kontralaterales und gehemmt durch ipsilaterales Ohr) als auch E0-Einheiten (erregt nur durch das kontralaterale Ohr).
- Neuronen sind tonotopisch organisiert und hochfrequent.
Anterolateraler periolivärer Nukleus (ALPO)
- Der Nukleus projiziert auf beide IC, wobei keine Zellen bilateral projizieren.
- Große multipolare Zellen projizieren sowohl bei der Katze als auch beim Meerschweinchen in den Nucleus cochlearis, aber nicht in den IC.
Ventromedialer periolivarer Nukleus (VMPO)
- Zwischen MSO und MNTB gelegen.
- Sendet Projektionen zum ICC auf beiden Seiten.
- Der Nukleus projiziert in beide IC, keine Zellen projizieren bilateral.
Rostraler periolivärer Nukleus (RPO) (Anteriorer periolivärer Nukleus (APO))
- befindet sich zwischen dem rostralen Pol des MSO und dem VNLL
Caudaler periolivarer Kern (CPO) (Posteriorer periolivarer Kern (PPO))
- Liegt zwischen dem kaudalen Pol des MSO und dem Facialis-Kern (7N)
Posteroventraler periolivarer Kern (PVPO)
- Der Kern projiziert in beide IC, wobei keine Zellen bilateral projizieren.
Pathophysiologie
Bei der Autopsie einer 21-jährigen Frau mit Autismus, Epilepsie und mentaler Retardierung wurde ein fast vollständiges Fehlen der oberen Olive festgestellt.
Siehe auch
- Olivarkörper
- Kulesza RJ, Cytoarchitecture of the human superior olivary complex: Medial and lateral superior olive. Hearing Research 225(2007) 80-90
- 2.0 2.1 Oliver DL, et al. Axonal projections from the lateral and medial superior olive to the inferior colliculus of the cat: a study using electron microscopic autoradiography. J Comp Neurol. 1995 Sep 11;360(1):17-32 Zitierfehler: Invalid
<ref>tag; name „R4“ defined multiple times with different content - Cajal, S. R. Y. and L. Azoulay (1909). Histologie du système nerveux de l’homme et des vertébrés. Paris, Maloine.
- 4.0 4.1 Kandel, et al Principles of Neuroscience. Fourth ed. pp 591-624. Copyright 2000, by McGraw-Hill Co. Zitierfehler: Ungültiger
<ref>-Tag; Name „R2“ mehrfach mit unterschiedlichem Inhalt definiert - Kulesza, R. J., Jr. (2007). „Cytoarchitecture of the human superior olivary complex: medial and lateral superior olive“. Hear Res 225(1-2): 80-90.
- Tsuchitani, C. und J. C. Boudreau (1967). „Encoding of stimulus frequency and intensity by cat superior olive S-segment cells“. J Acoust Soc Am 42(4): 794-805.
- 8.0 8.1 8.2 Zitierfehler: Invalid
<ref>tag; name „R10“ defined multiple times with different content - Wu, S. H. and J. B. Kelly (1994). „Physiologische Beweise für ipsilaterale Hemmung in der lateralen superioren Olive: synaptische Antworten in Mäusegehirnschnitten.“ Hear Res 73(1): 57-64.
- Brownell, W. E., P. B. Manis, et al. (1979). „Ipsilaterale inhibitorische Antworten in der lateralen superioren Olive der Katze“. Brain Res 177(1): 189-93.
- Darrow, K. N., S. F. Maison, et al. (2006). „Cochlear efferent feedback balances interaural sensitivity.“ Nat Neurosci 9(12): 1474-6.
- Warr, W. B. und J. J. Guinan, Jr. (1979). „Efferente Innervation des Cortischen Organs: zwei getrennte Systeme“. Brain Res 173(1): 152-5.
- 13.0 13.1 Schofield, B. R. und N. B. Cant (1991). „Organization of the superior olivary complex in the guinea pig. I. Cytoarchitecture, cytochrome oxidase histochemistry, and dendritic morphology.“ J Comp Neurol 314(4): 645-70. Zitierfehler: Invalid
<ref>tag; name „R7“ defined multiple times with different content - 14.0 14.1 14.2 14.3 14.4 14.5 14.6 14.7 14.8 Illing, R. B., K. S. Kraus, et al. (2000). „Plastizität des superioren Olivenkomplexes“. Microsc Res Tech 51(4): 364-81. Zitierfehler: Ungültiger
<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Zitierfehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Zitierfehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Zitierfehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R5“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Invalid<ref>tag; name „R5“ defined multiple times with different content - Warr, W. B. and J. E. Beck (1996). „Multiple Projektionen aus dem ventralen Nukleus des Trapezkörpers bei der Ratte“. Hear Res 93(1-2): 83-101.
- 16.00 16.01 16.02 16.03 16.04 16.05 16.06 16.07 16.08 16.09 16.10 Schofield, B. R. und N. B. Cant (1992). „Organisation des superioren olivären Komplexes beim Meerschweinchen: II. Muster der Projektion von den periolivären Kernen zum Colliculus inferior“. J Comp Neurol 317(4): 438-55. Cite error: Ungültiger
<ref>-Tag; Name „R6“ mehrfach definiert mit unterschiedlichem Inhalt Zitierfehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Zitierfehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Zitierfehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Ungültiger<ref>-Tag; Name „R6“ mehrfach mit unterschiedlichem Inhalt definiert Cite-Fehler: Invalid<ref>tag; name „R6“ defined multiple times with different content - Smith, P. H., P. X. Joris, et al. (1993). „Projektionen physiologisch charakterisierter sphärischer Buschzellaxone aus dem Cochlea-Kern der Katze: Nachweis von Verzögerungslinien zur medialen superioren Olive.“ J Comp Neurol 331(2): 245-60.
- Wenthold, R. J., D. Huie, et al. (1987). „Glycin-Immunreaktivität lokalisiert im Nucleus cochlearis und im superioren olivären Komplex“. Neuroscience 22(3): 897-912.
- 19.0 19.1 Adams, J. C. (1983). „Cytology of periolivary cells and the organization of their projections in the cat.“ J Comp Neurol 215(3): 275-89. Cite error: Invalid
<ref>tag; name „R8“ defined multiple times with different content - Friauf, E. and J. Ostwald (1988). „Divergente Projektionen von physiologisch charakterisierten Neuronen des ventralen Nucleus cochlearis der Ratte, nachgewiesen durch intraaxonale Injektion von Meerrettichperoxidase.“ Exp Brain Res 73(2): 263-84.
- Kulesza, R. J., Jr. und A. S. Berrebi (2000). „Der Nucleus superior paraolivaris der Ratte ist ein GABA-erger Kern“. J Assoc Res Otolaryngol 1(4): 255-69.
- Kulesza, R. J., Jr., G. A. Spirou, et al. (2003). „Physiologische Reaktionseigenschaften von Neuronen im Nucleus paraolivaris superior der Ratte“. J Neurophysiol 89(4): 2299-312.
- WHITE JS, WARR WB. Der doppelte Ursprung des olivocochleären Bündels bei der Albino-Ratte. J. Comp. Neurol. 219:203-214, 1983.
- Guinan, J. J., Jr., S. S. Guinan, et al. (1972). „Single auditory units in the superior olivary complex. I. Responses to sounds and classifications based on physiological properties.“ Int J Neurosci 4: 101-20.
- Tsuchitani, C. (1977). „Funktionelle Organisation der lateralen Zellgruppen des superioren olivären Komplexes der Katze“. J Neurophysiol 40(2): 296-318.
- Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996). Embryologischer Ursprung des Autismus: Entwicklungsanomalien der motorischen Kerne des Hirnnervs. J Comp Neurol 370 (2): 247-61.
- MedEd at Loyola Neuro/frames/nlBSs/nl27fr.htm
- Image at med.utah.edu (pick 9b. Pons)
Dieser Artikel beruhte ursprünglich auf einem Eintrag aus einer gemeinfreien Ausgabe von Gray’s Anatomy. Daher können einige der darin enthaltenen Informationen veraltet sein. Bitte bearbeiten Sie den Artikel, wenn dies der Fall ist, und entfernen Sie diesen Hinweis, wenn er nicht mehr relevant ist.
|
v-d-e
Gehirn: Rhombencephalon (Hinterhirn) |
||||||
|---|---|---|---|---|---|---|
| Metencephalon/ Pons |
|
Dorsal/
(tegmentum) |
Ventral/
(Basis) |
Raphe/retikulär
|
||
| Myelencephalon/ Medulla |
|
Dorsal
|
Ventral
|
Raphe/retikuläre
|
||
|
v-d-e
Sinnessystem: Auditives und vestibuläres System (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Außenohr |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Ohrläppchen) – Gehörgang – Ohrmuschelmuskeln Ohrmuschel (Umbo, Pars flaccida) |
|||||||||||
| Mittelohr |
|
|||||||||||
| Innenohr/ (membranöses Labyrinth, knöchernes Labyrinth) |
|
|||||||||||
| {| class=“navbox collapsible nowraplinks“ style=“margin:auto;“ | ||||||||||||
|
—
|
||||||||||||
|
||||||||||||
|}
Diese Seite verwendet Creative Commons lizenzierte Inhalte aus Wikipedia (Autoren anzeigen).
Schreibe einen Kommentar